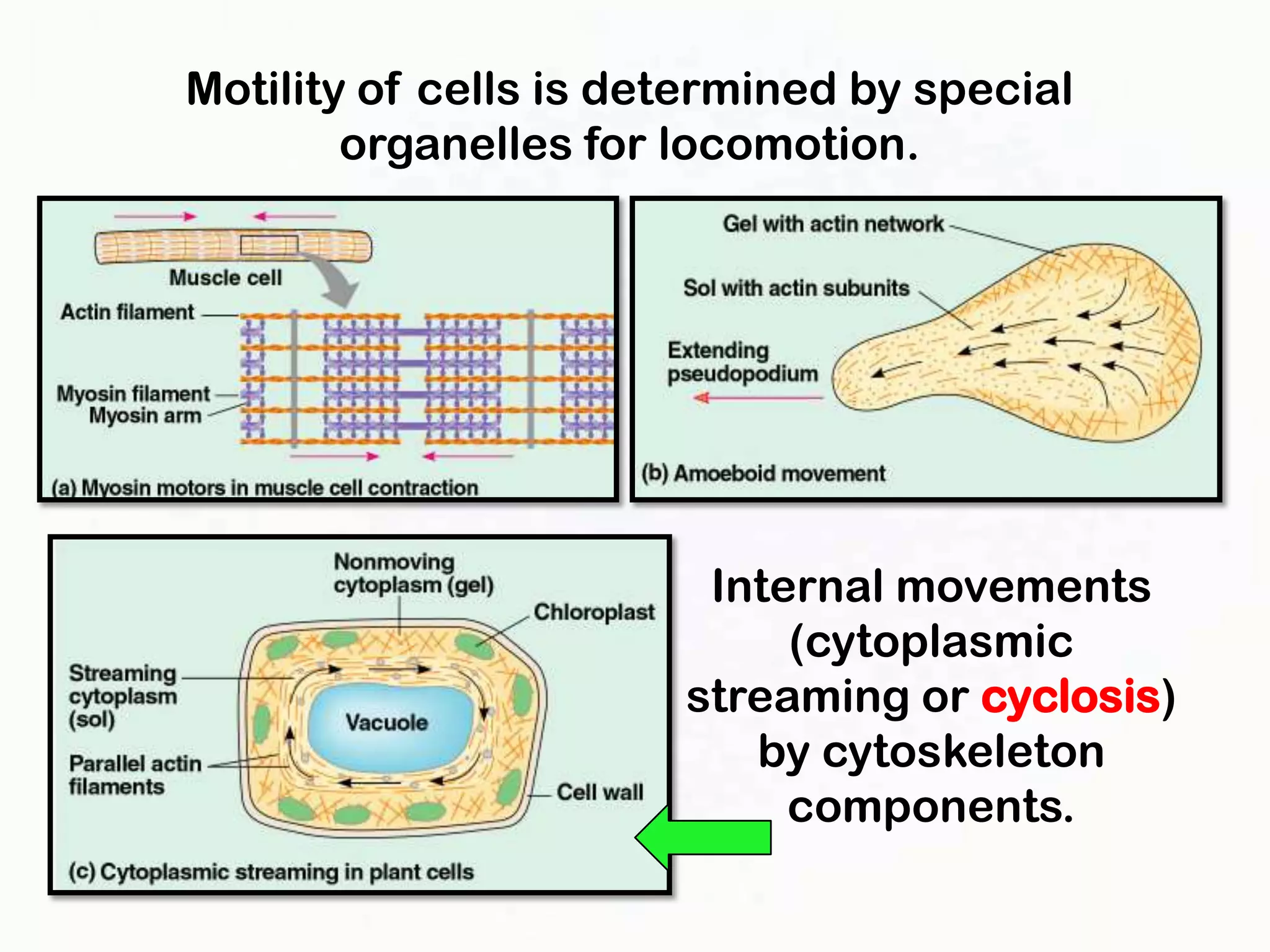
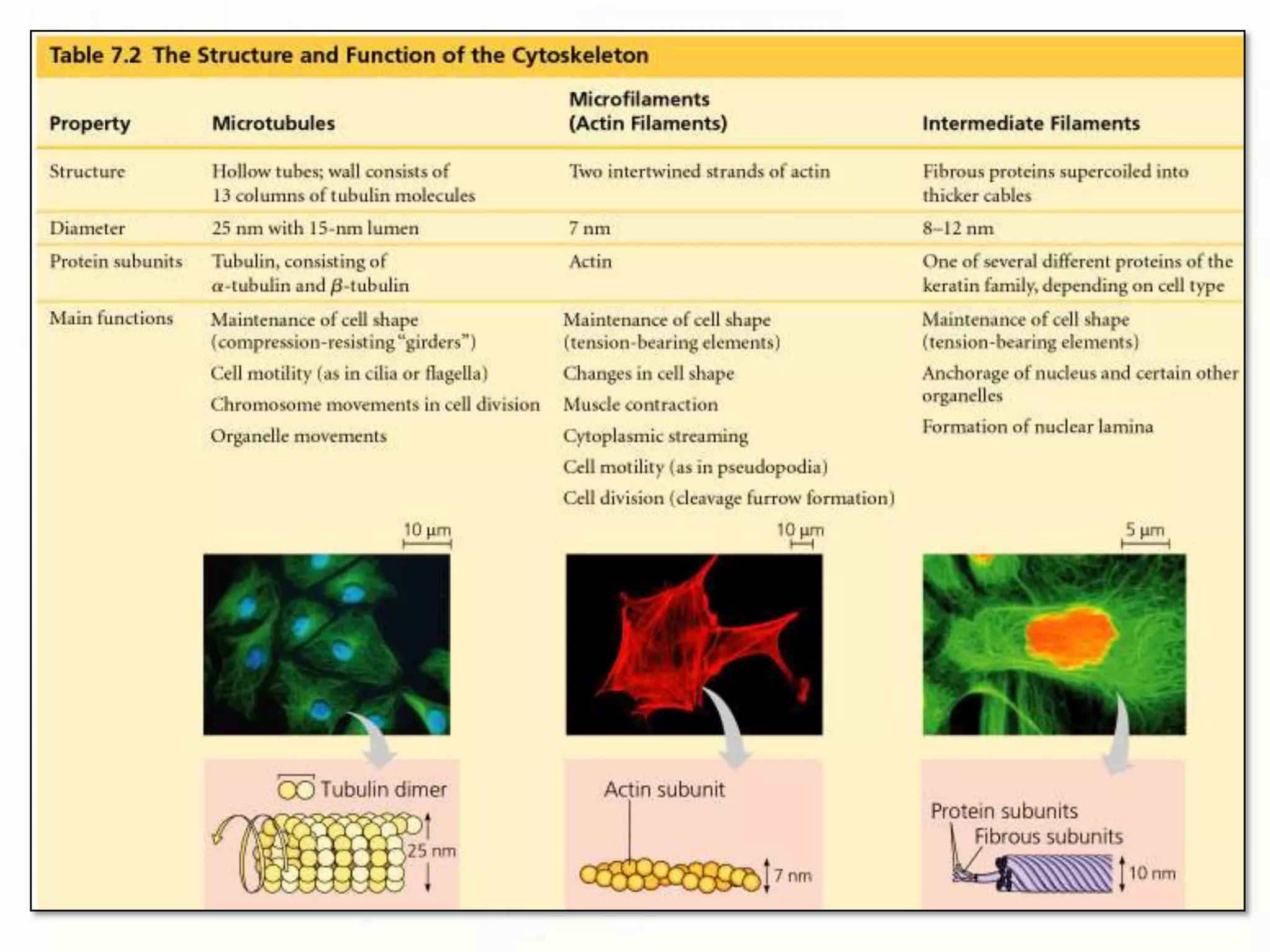
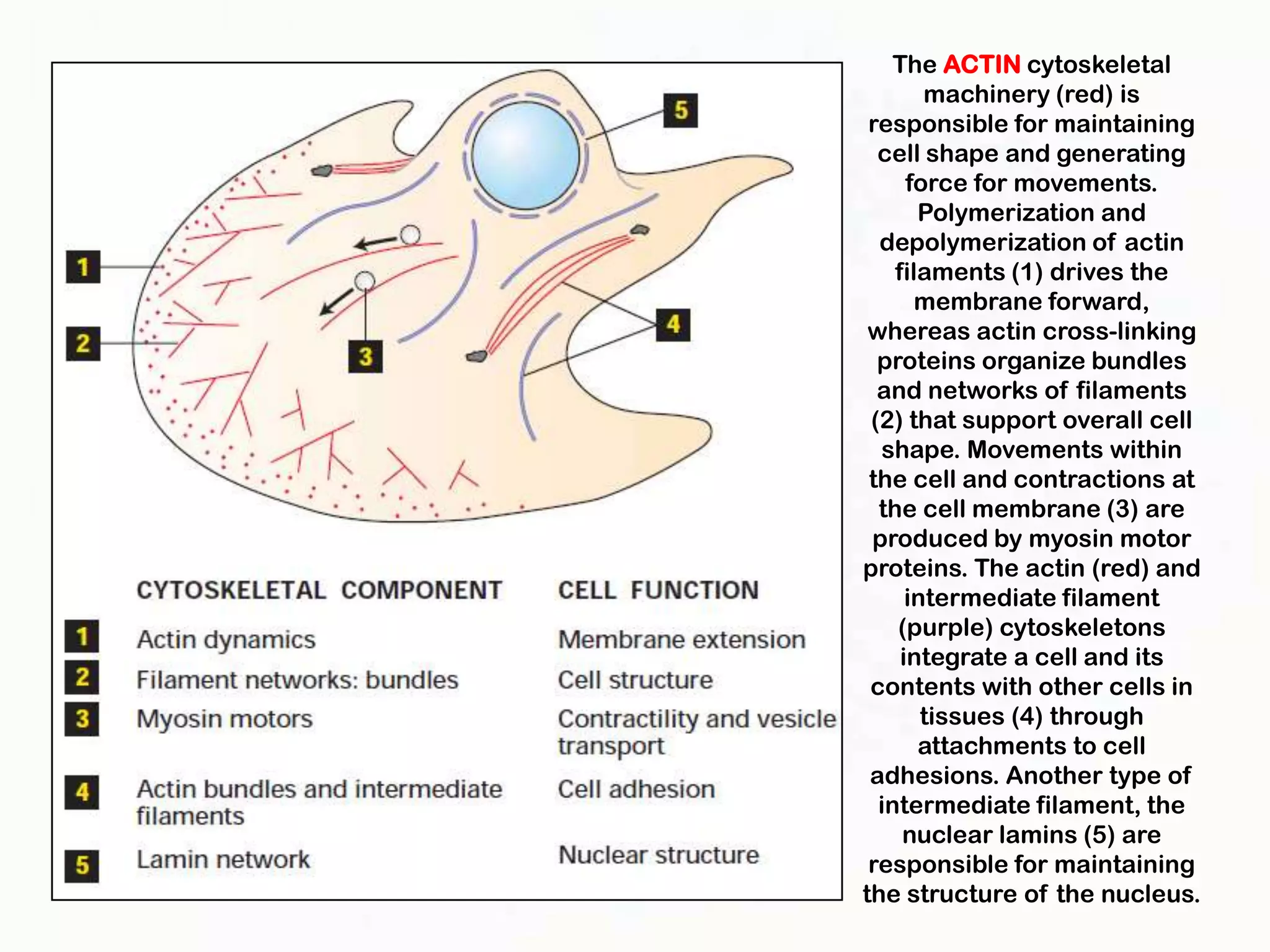
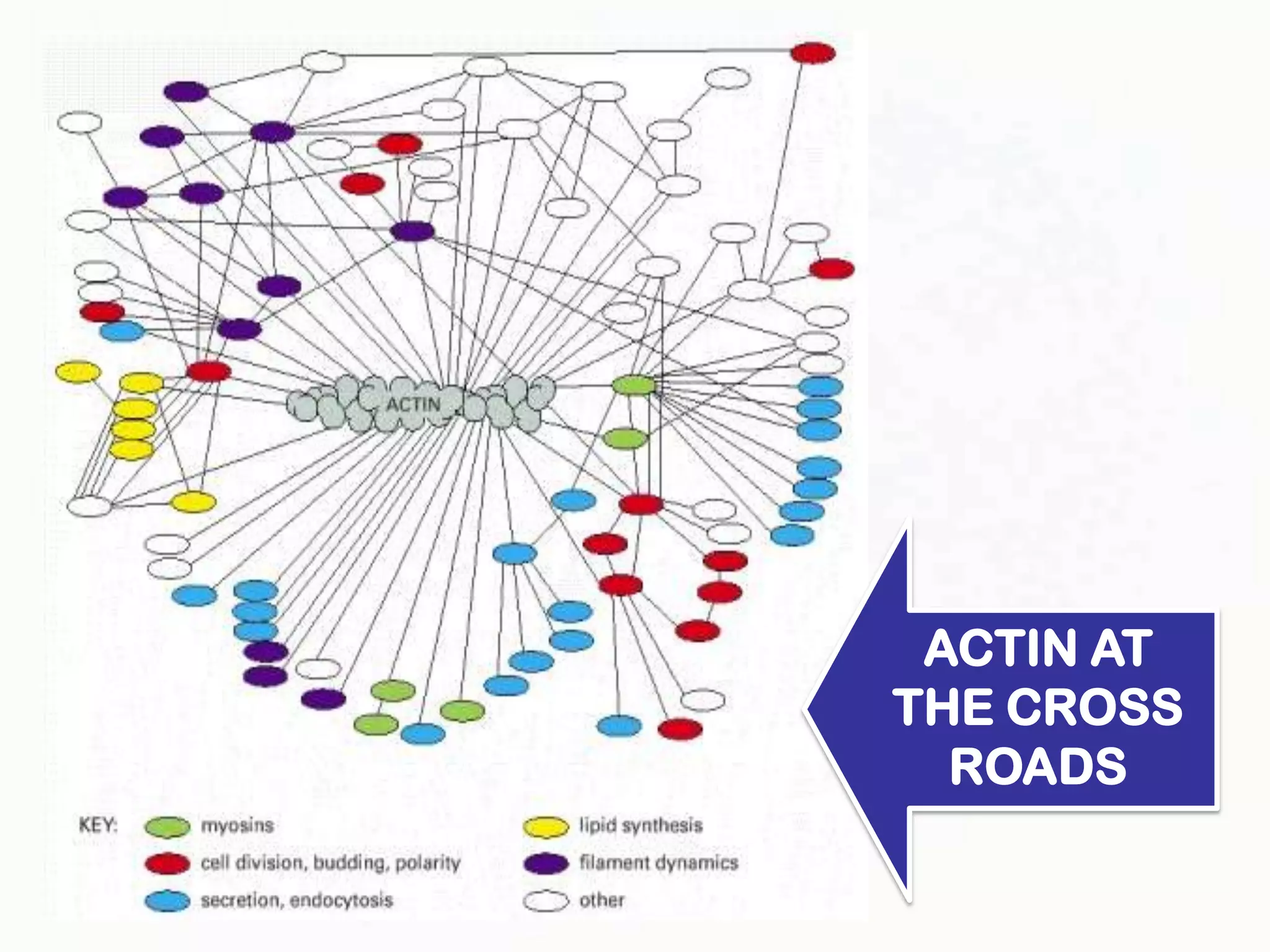
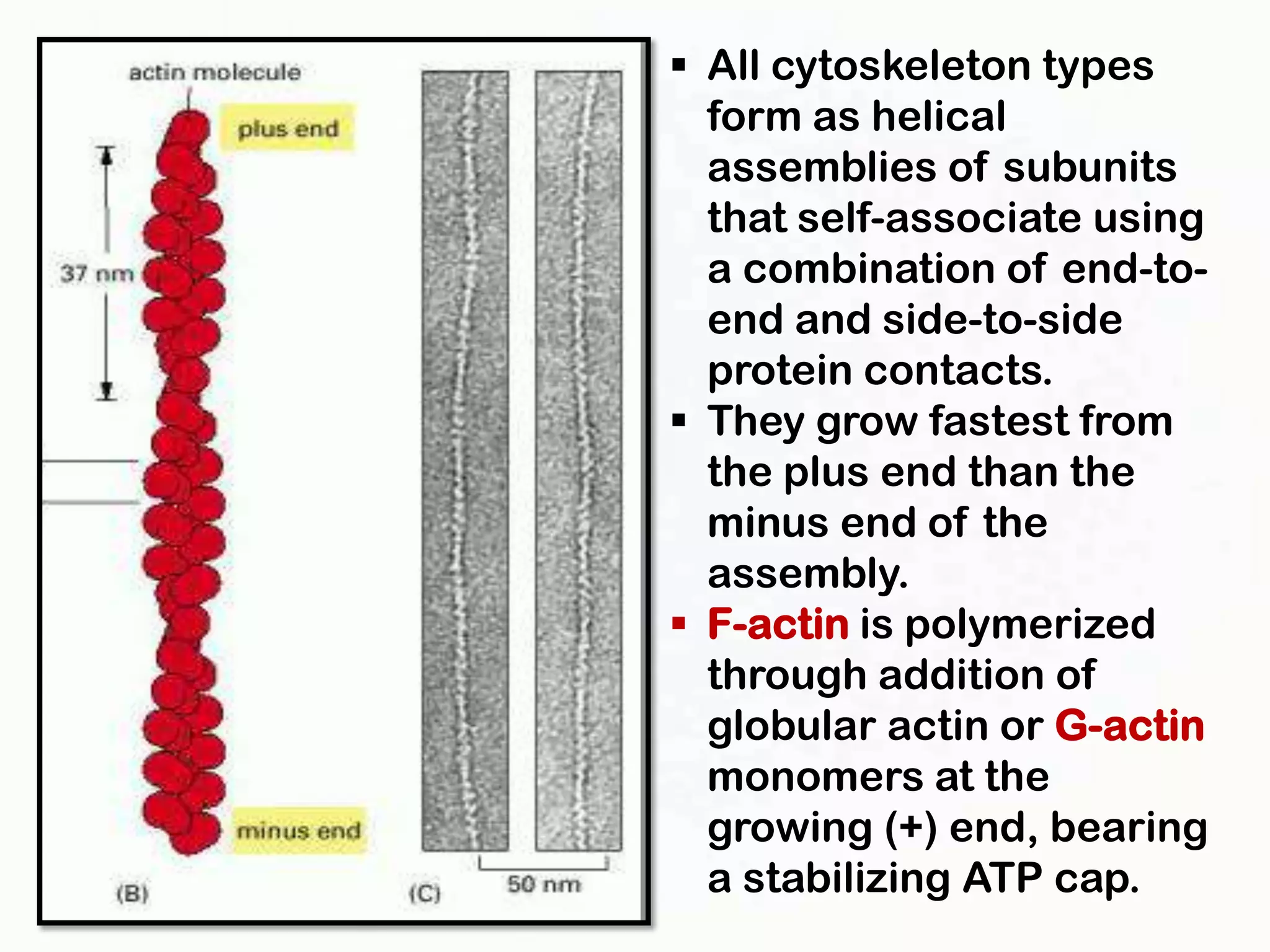
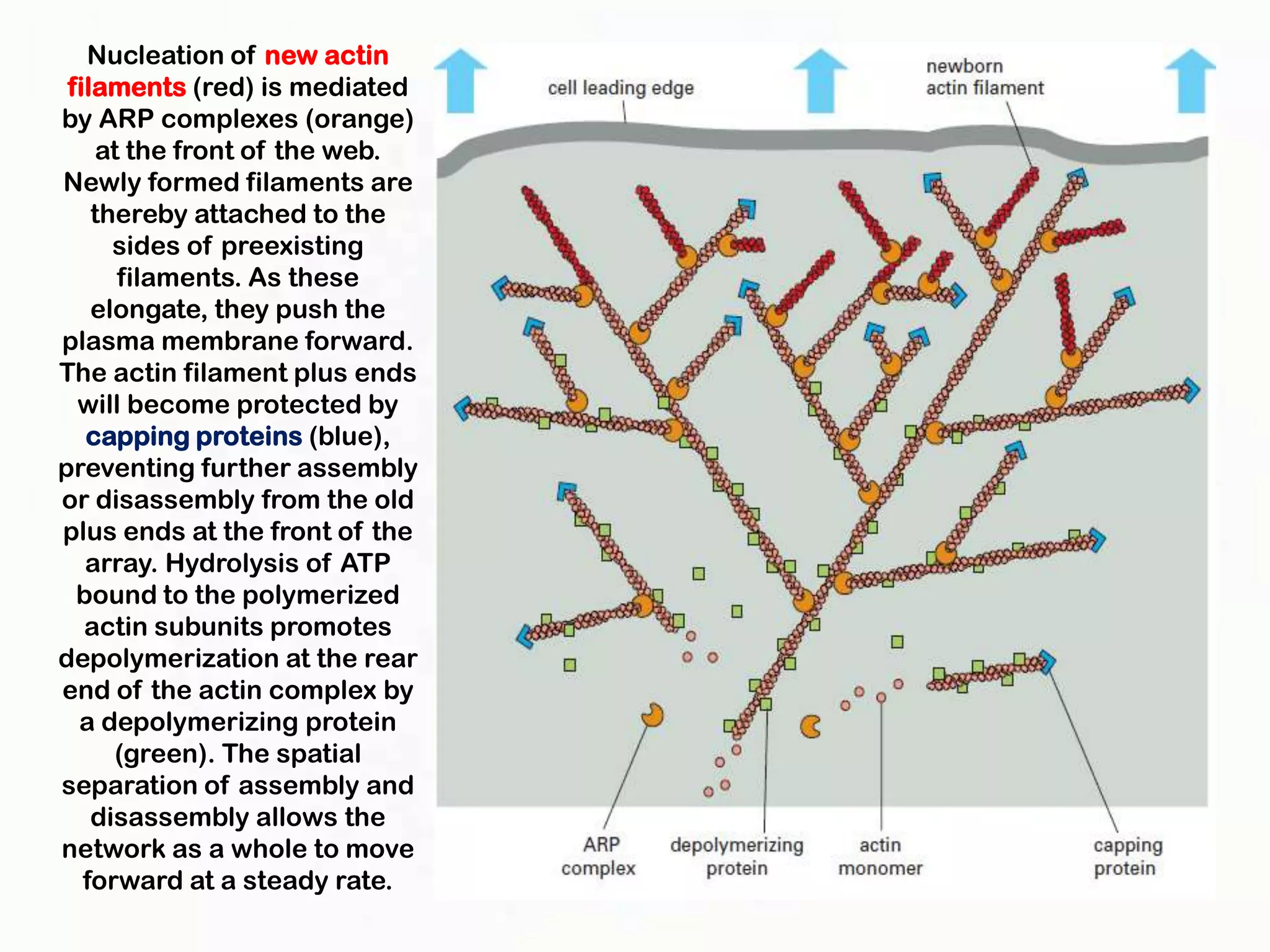
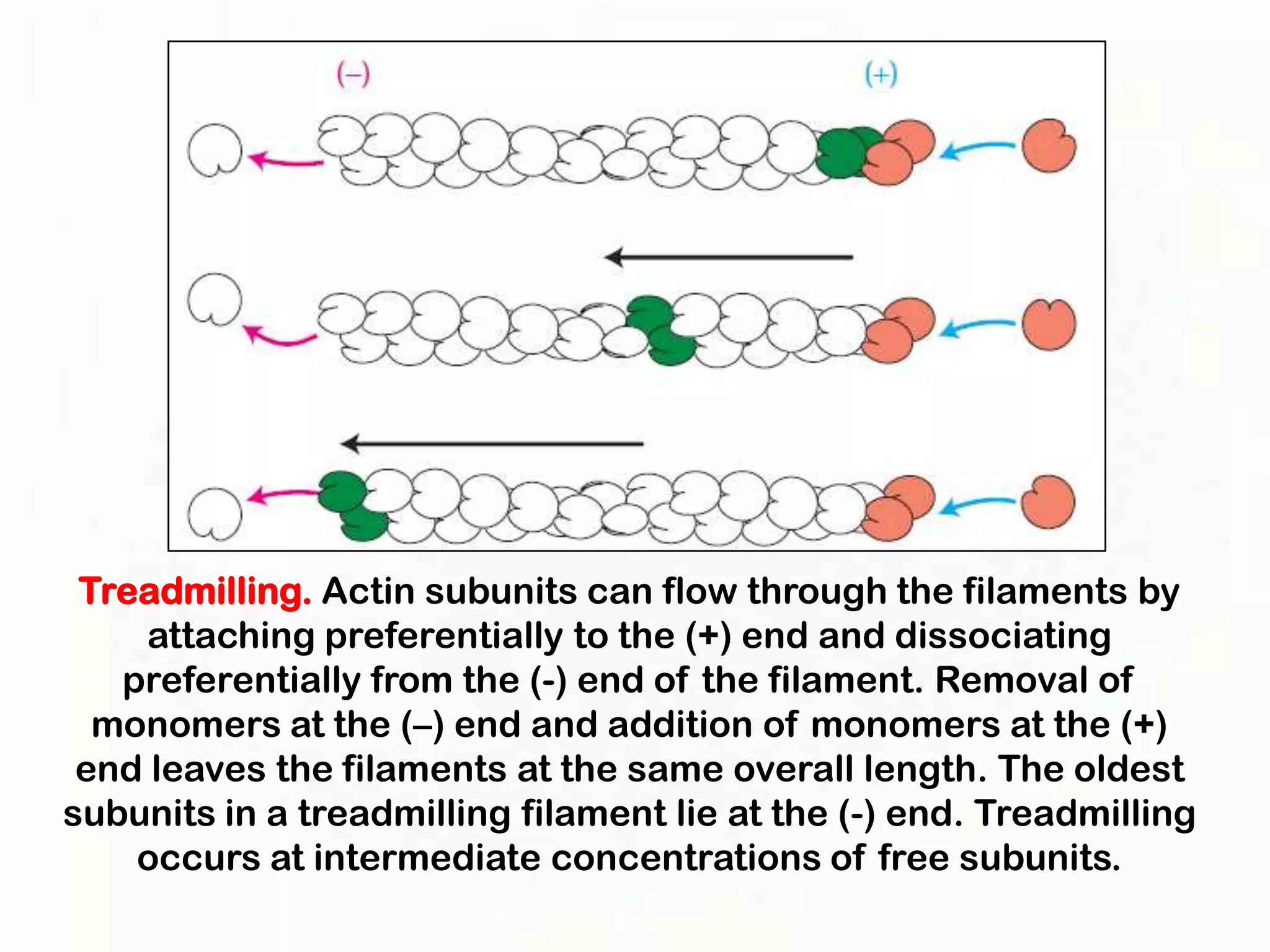
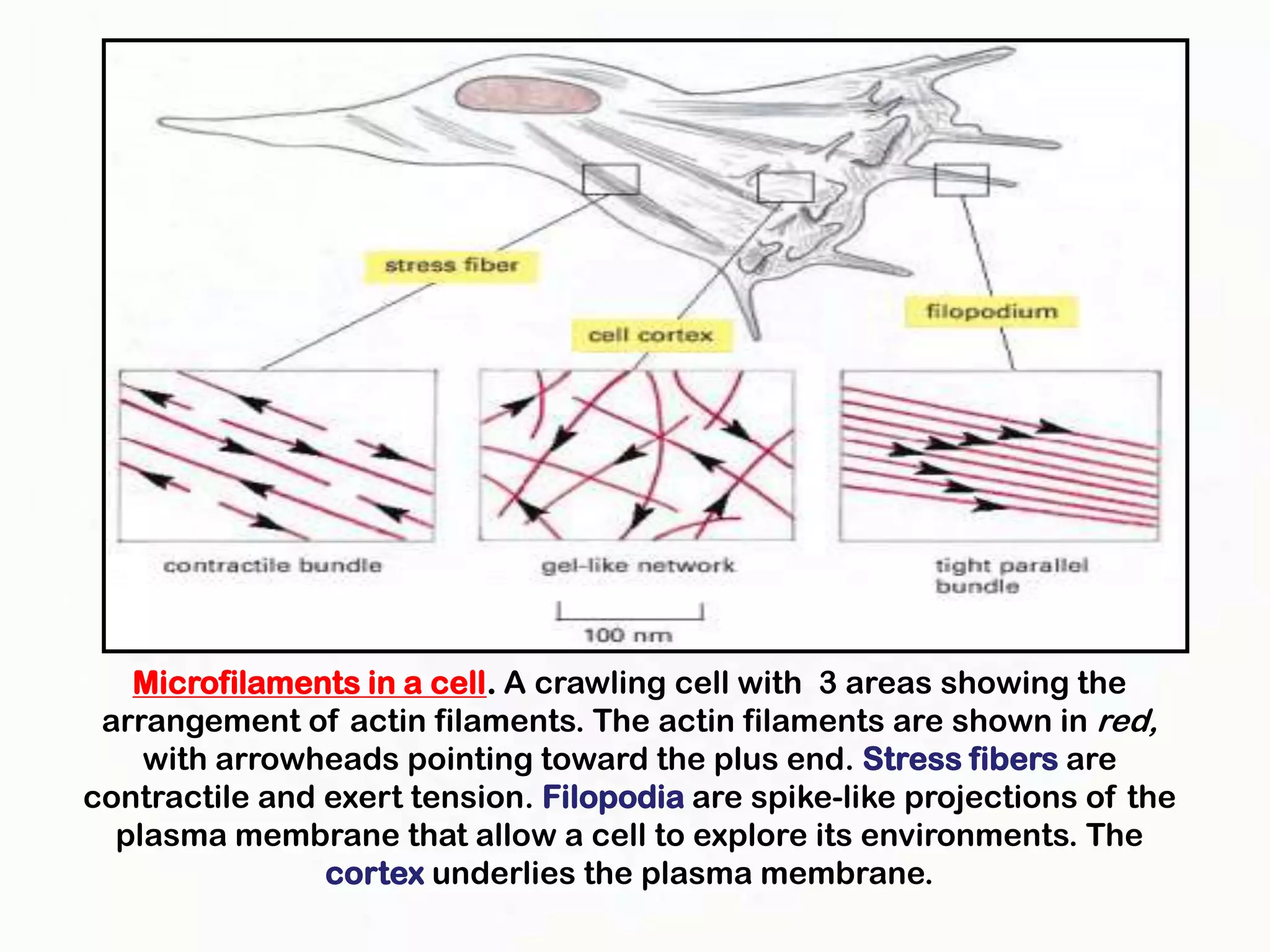
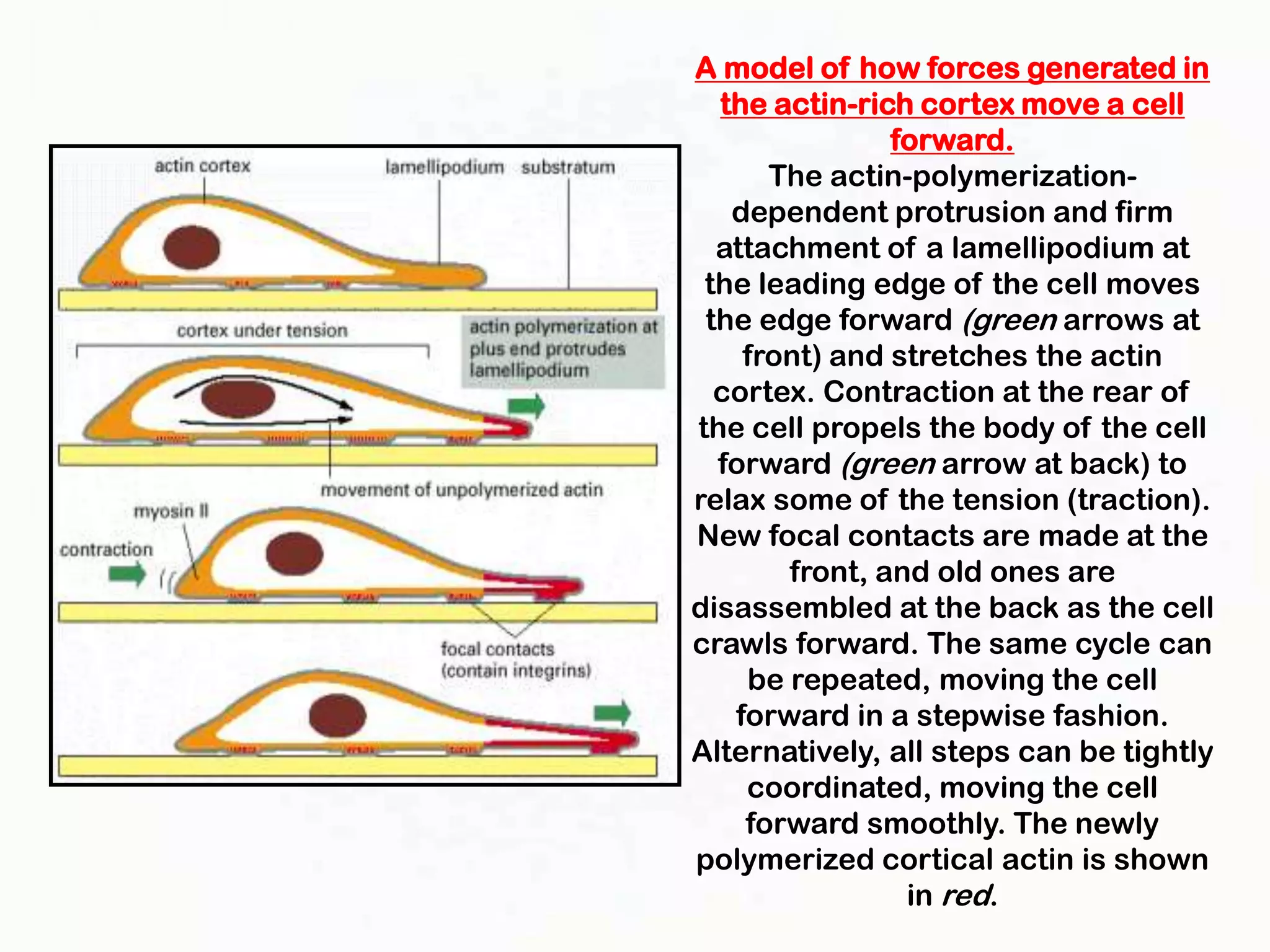
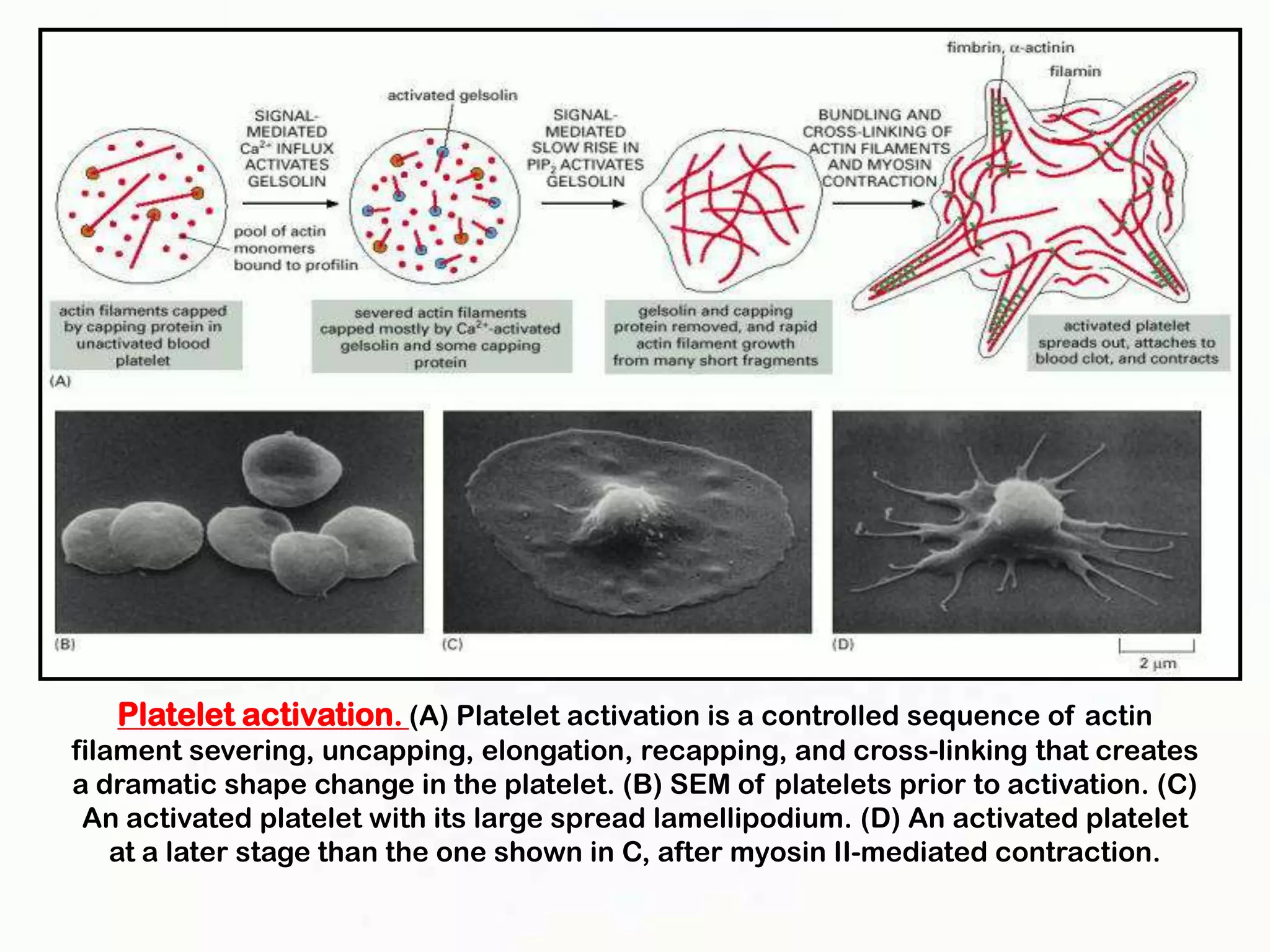
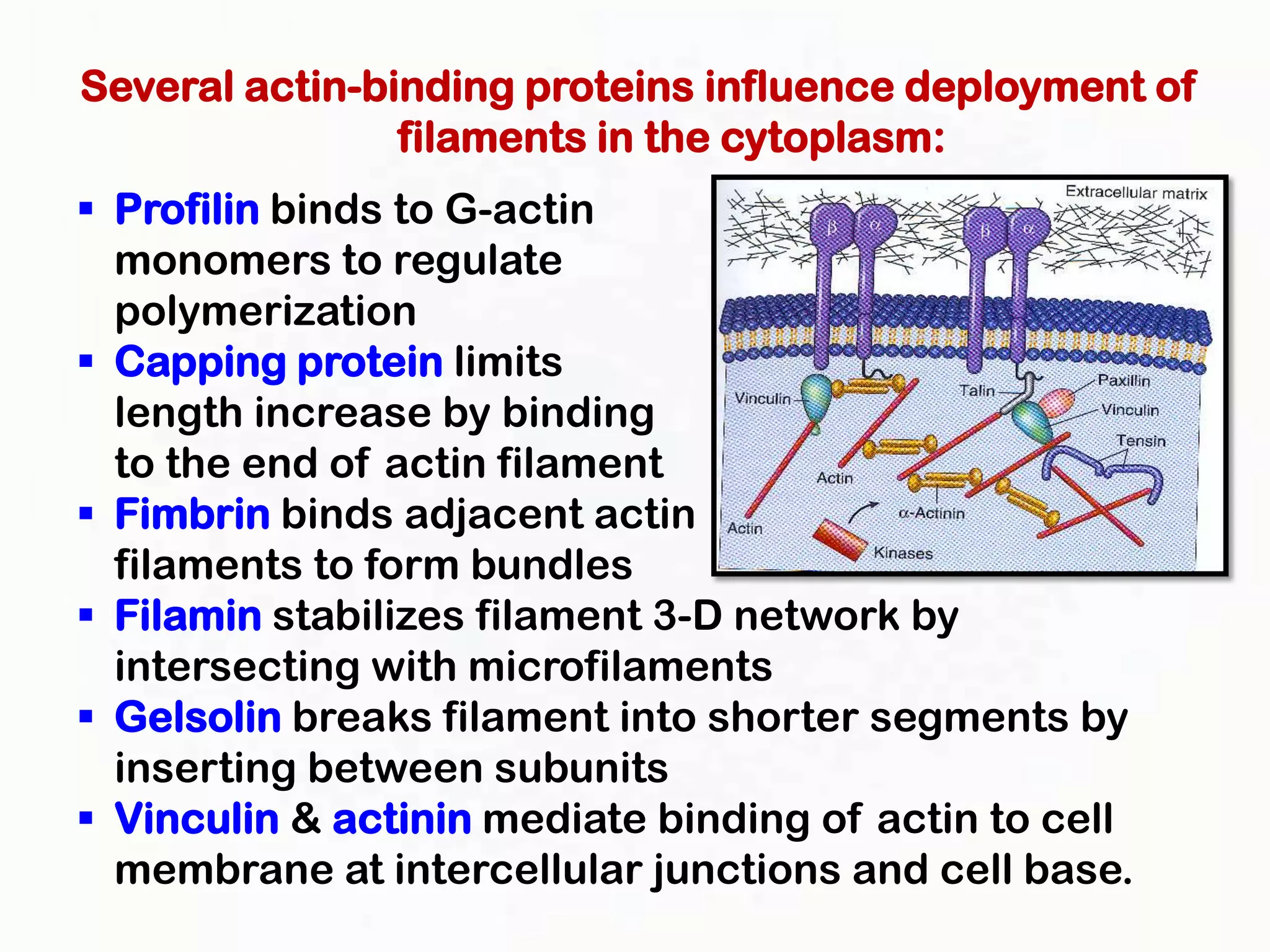
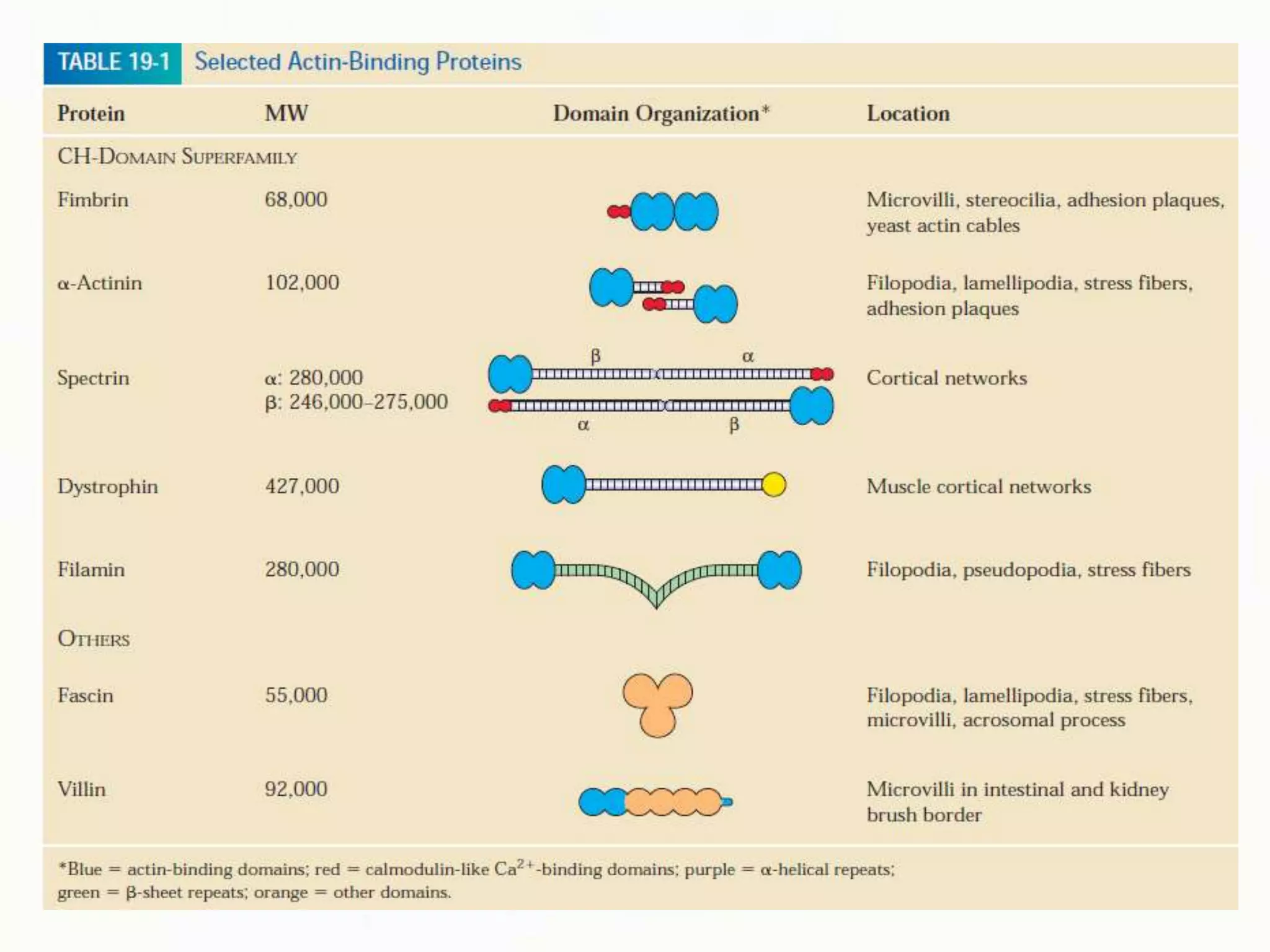
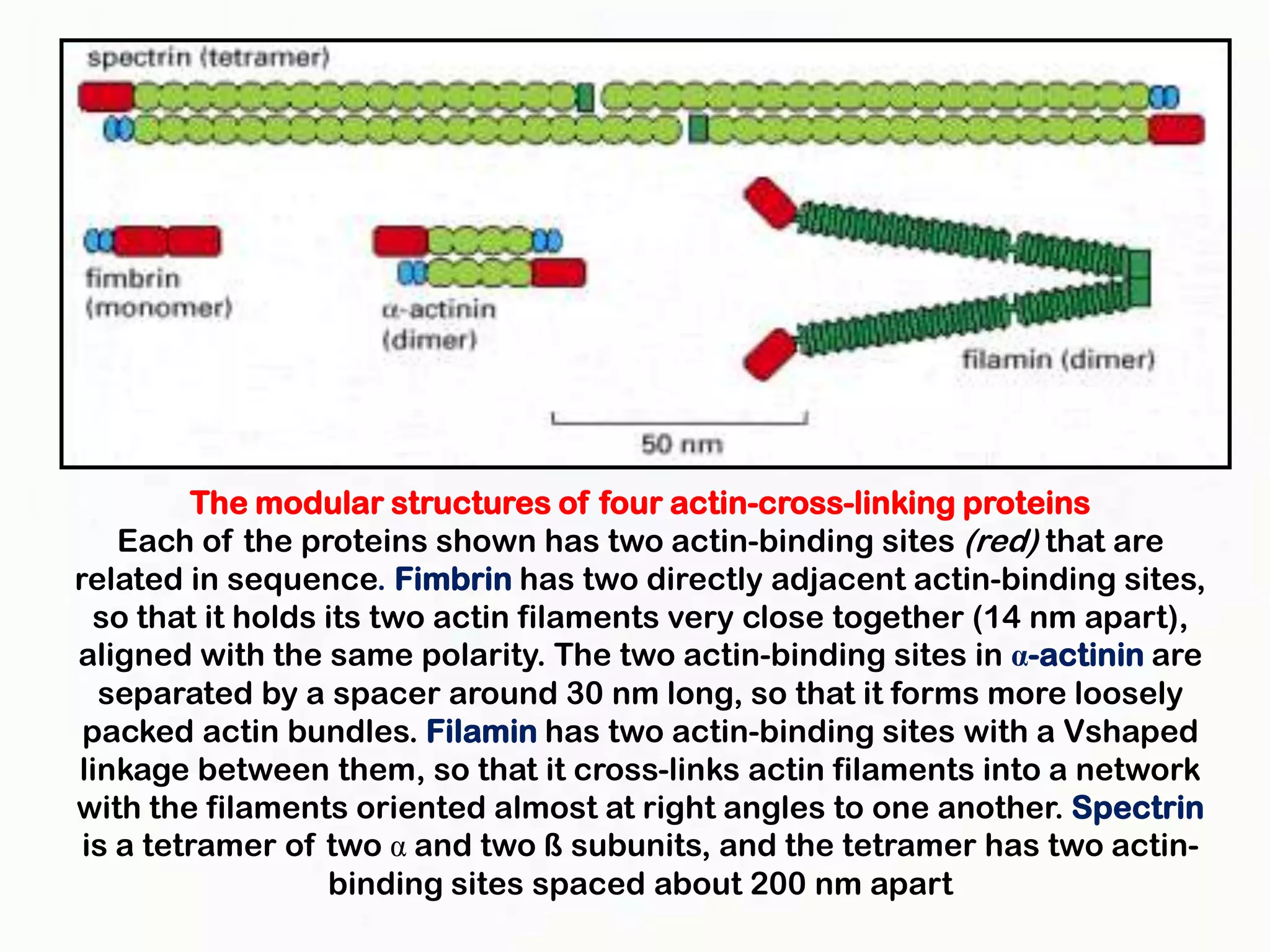
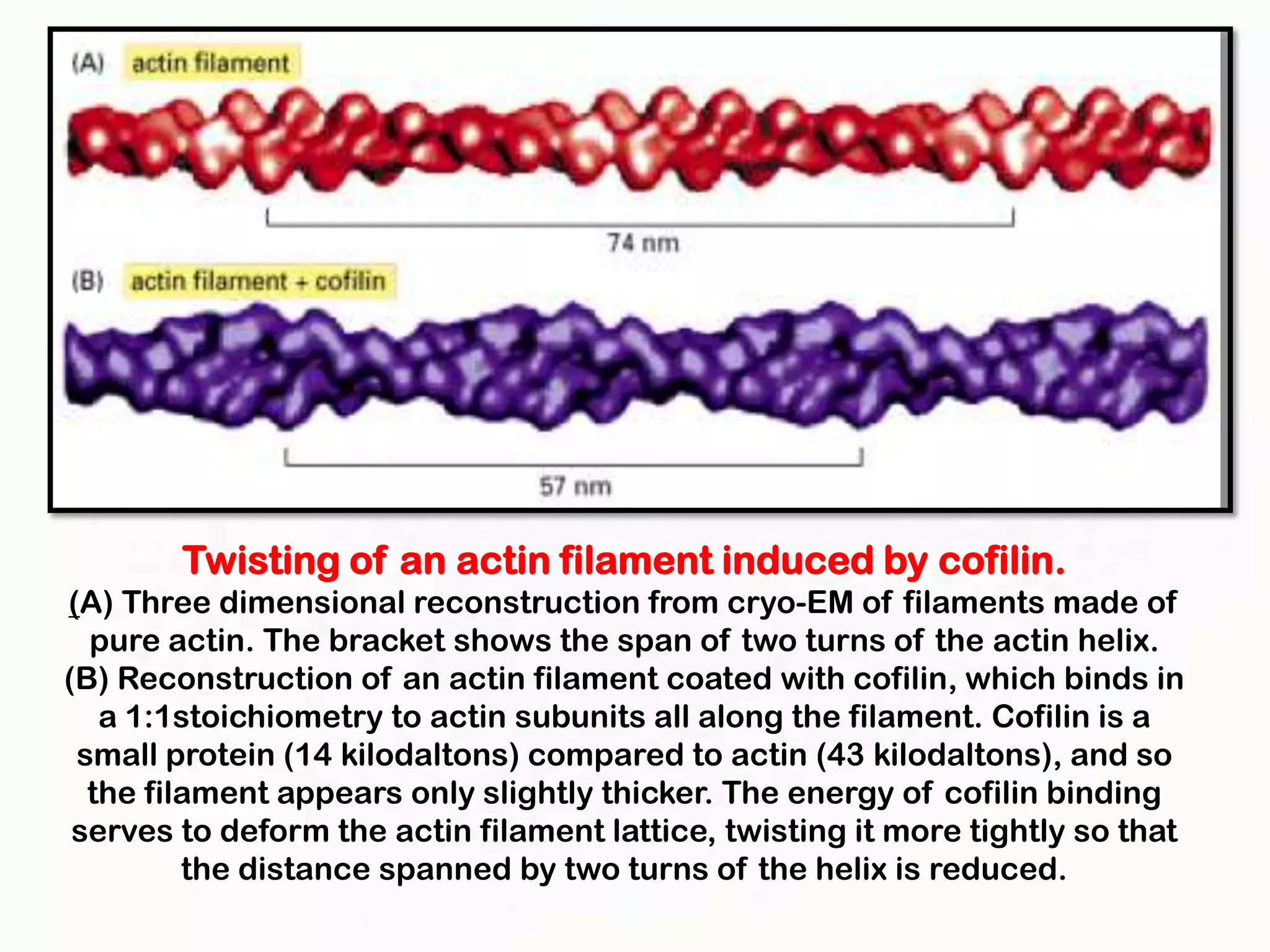
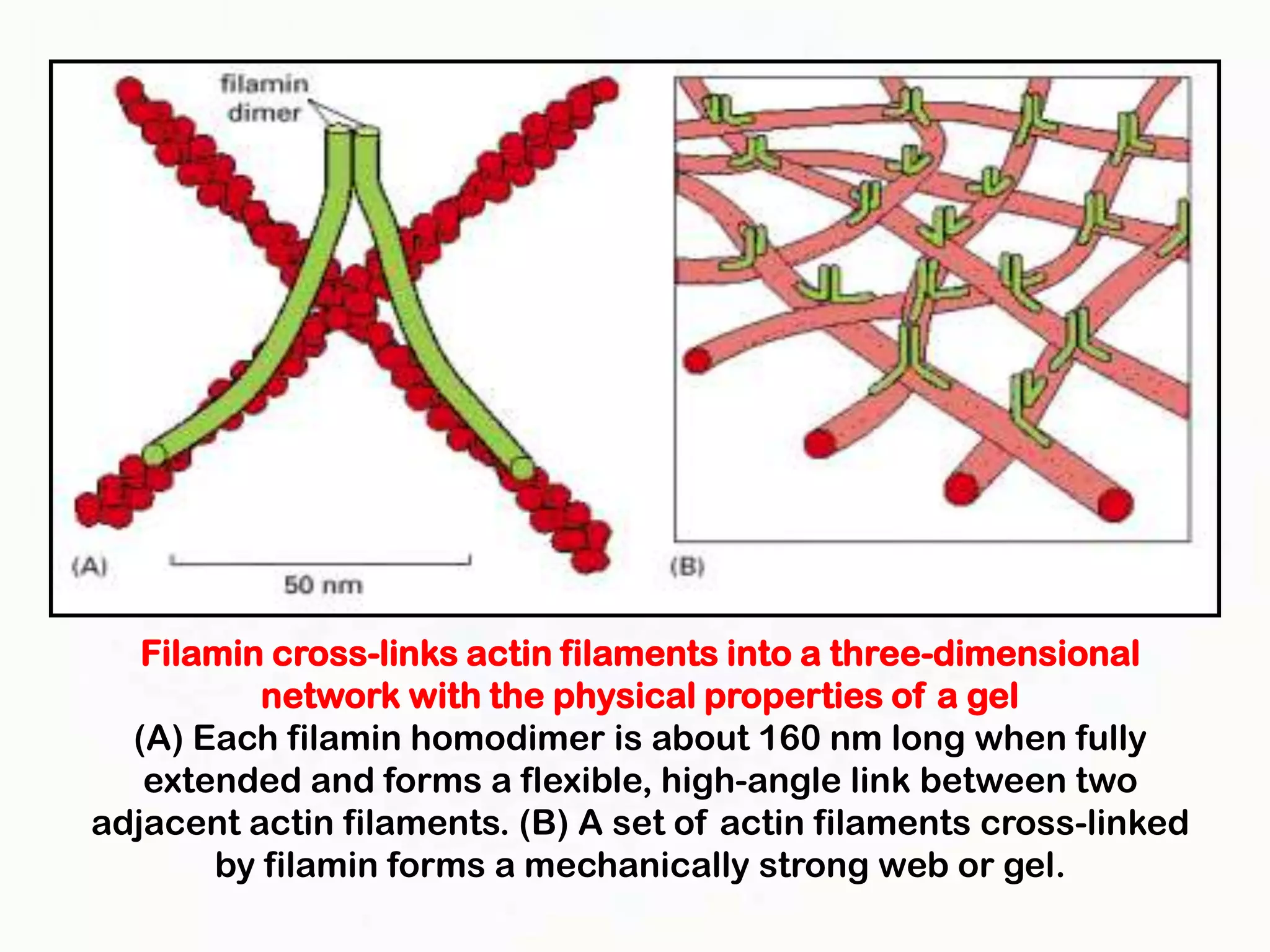
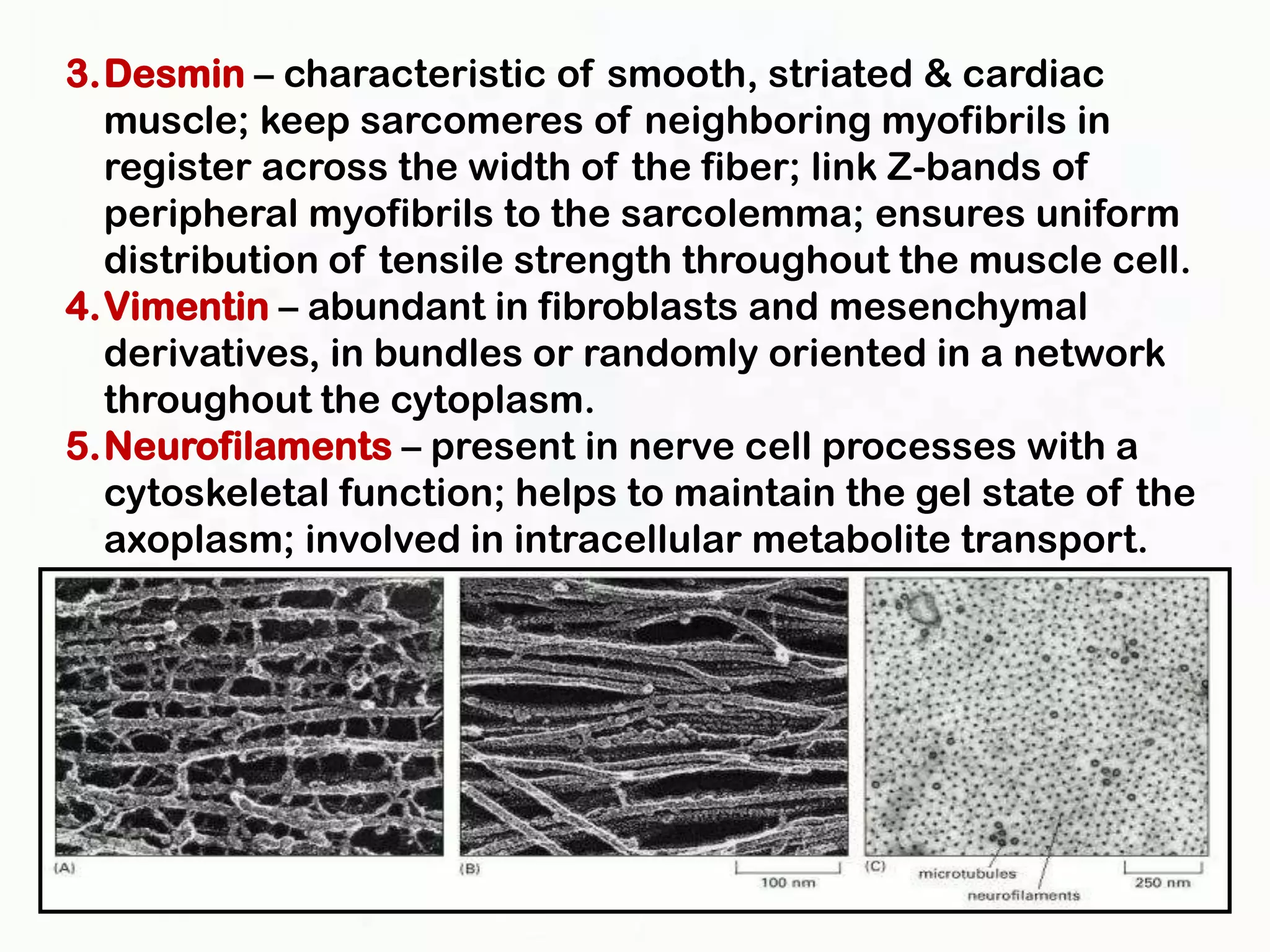
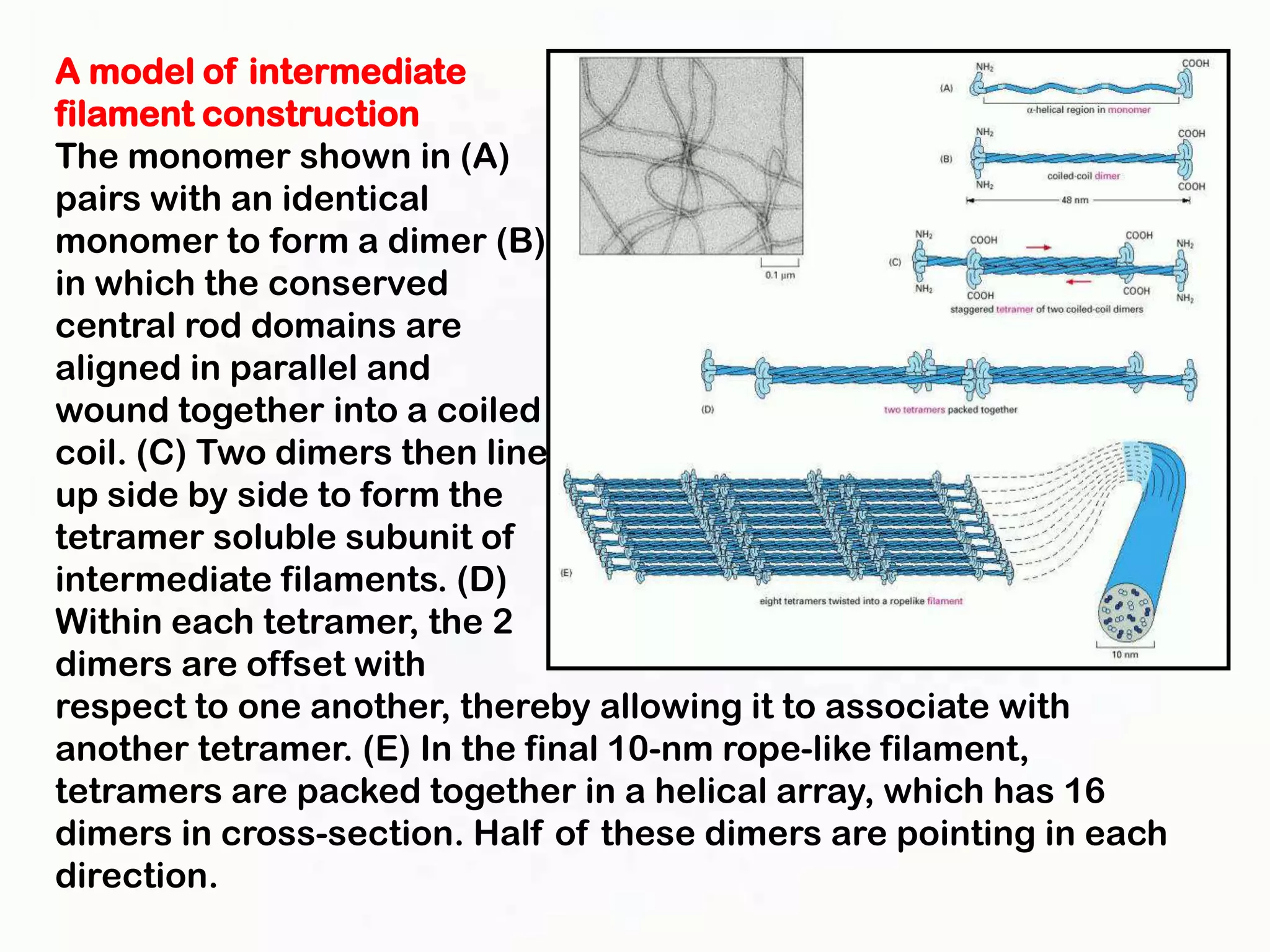
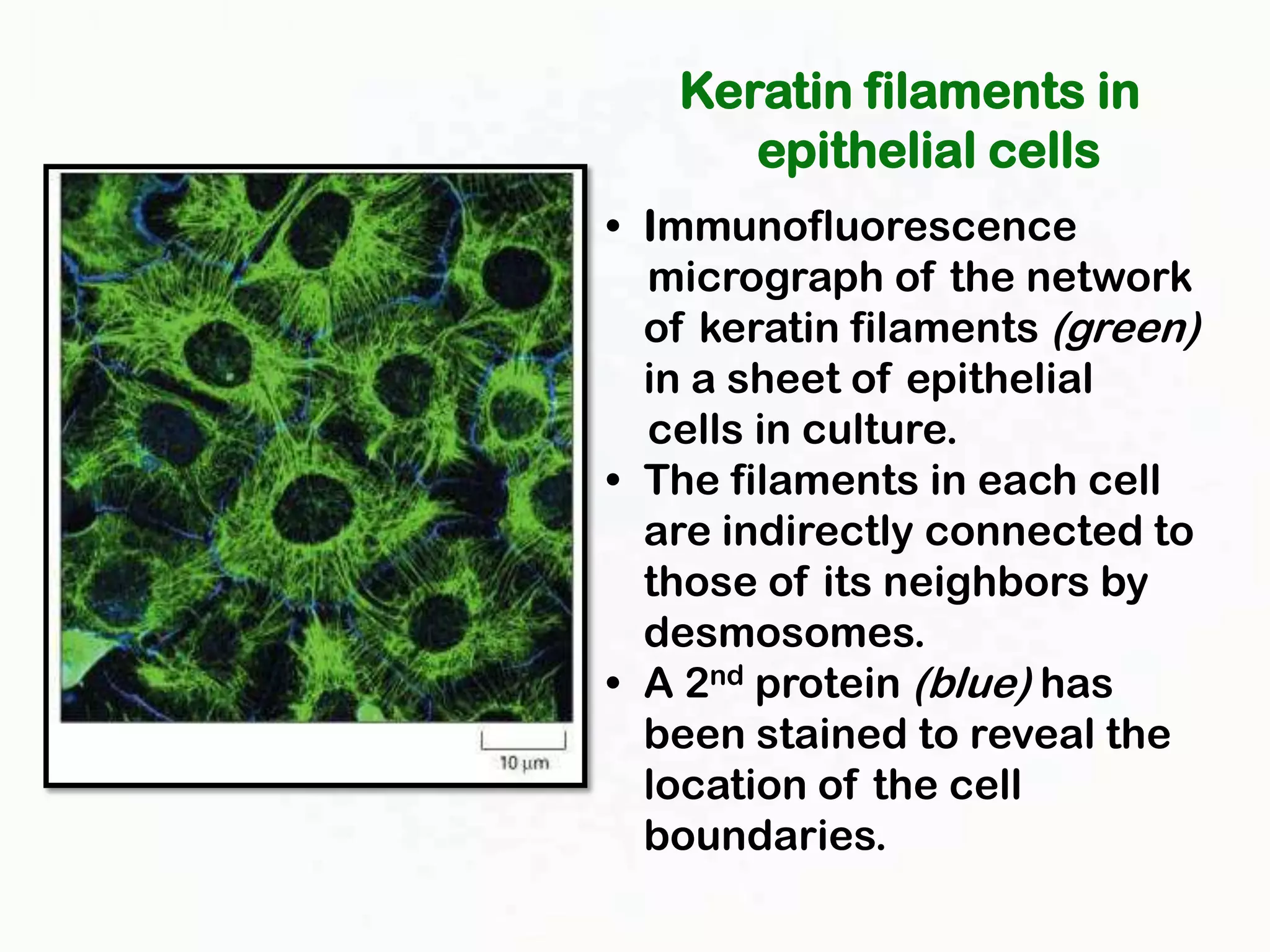
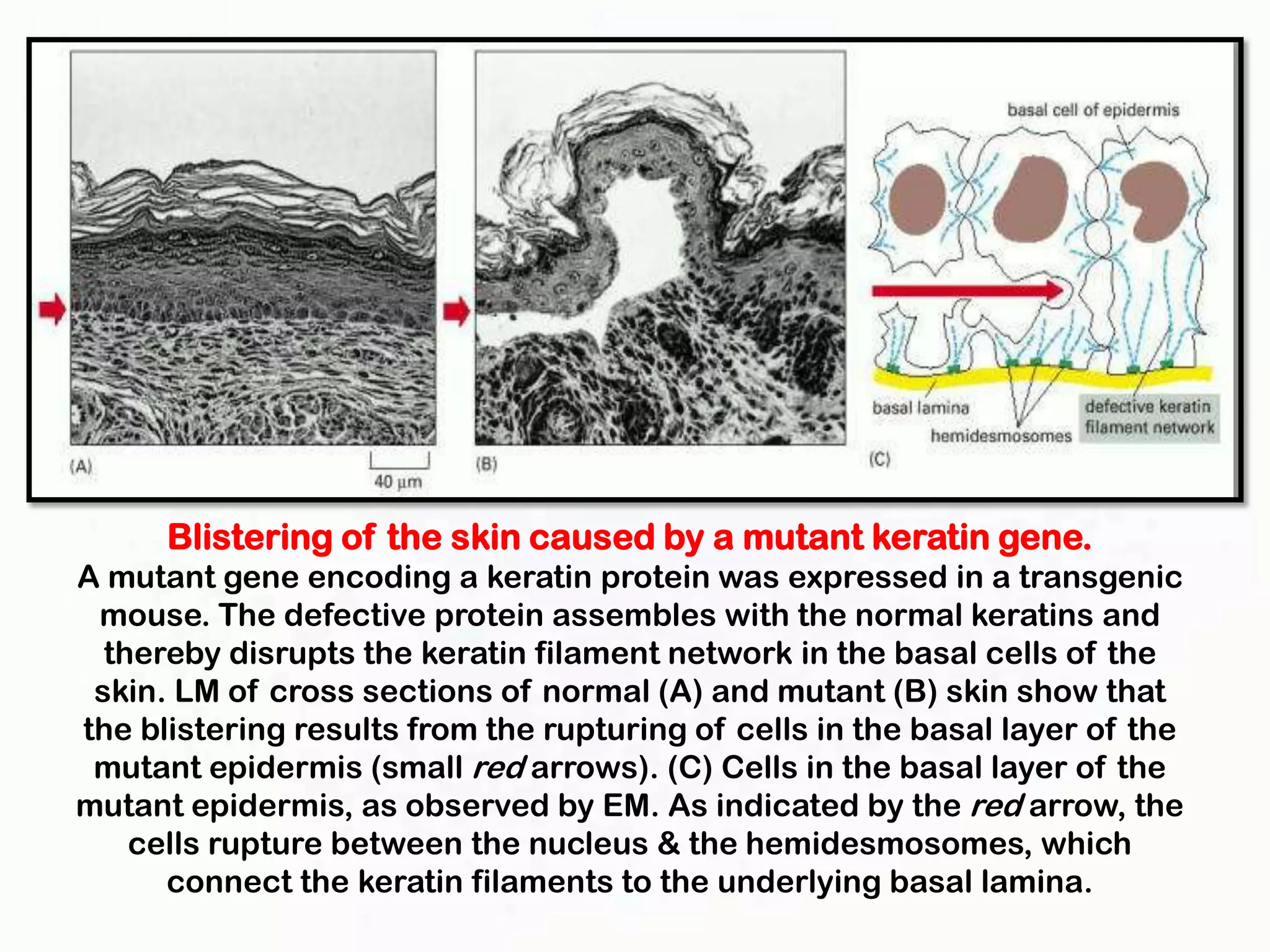
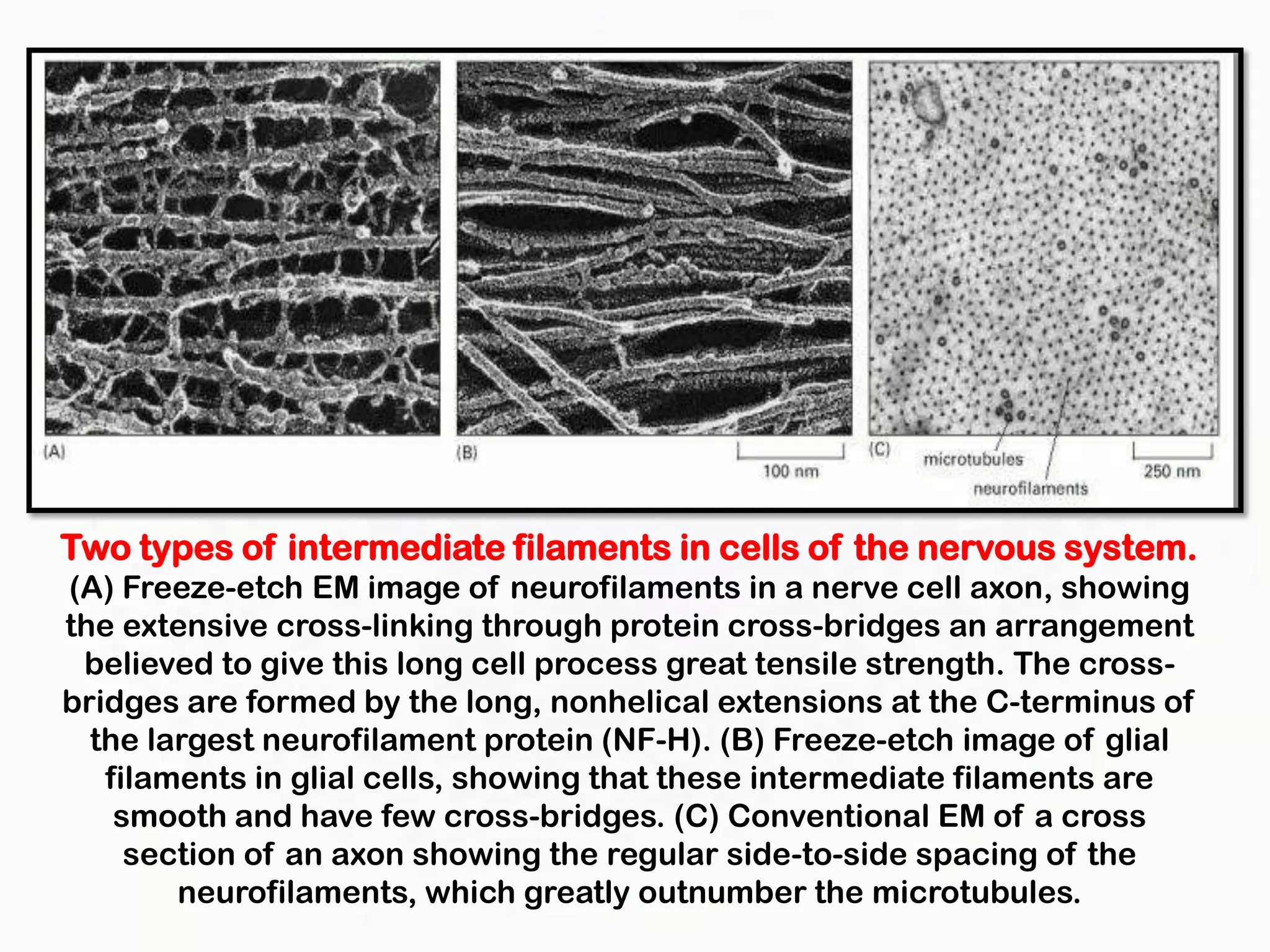
The document summarizes key aspects of the cytoskeleton, focusing on actin filaments. It describes how actin filaments: 1) maintain cell shape and generate force for cell movements through polymerization and depolymerization; 2) integrate cells through attachments to cell adhesions; and 3) produce movements within cells and at the cell membrane through interactions with myosin motor proteins. Accessory proteins regulate actin dynamics by capping, bundling, severing, and crosslinking filaments.
































































































![Vibe Coding vs. Spec-Driven Development [Free Meetup]](https://cdn.slidesharecdn.com/ss_thumbnails/vibecodingvsspecdrivendevelopment-251209105622-43f455e7-thumbnail.jpg?width=640&height=640&fit=bounds)













