Downloaded 109 times

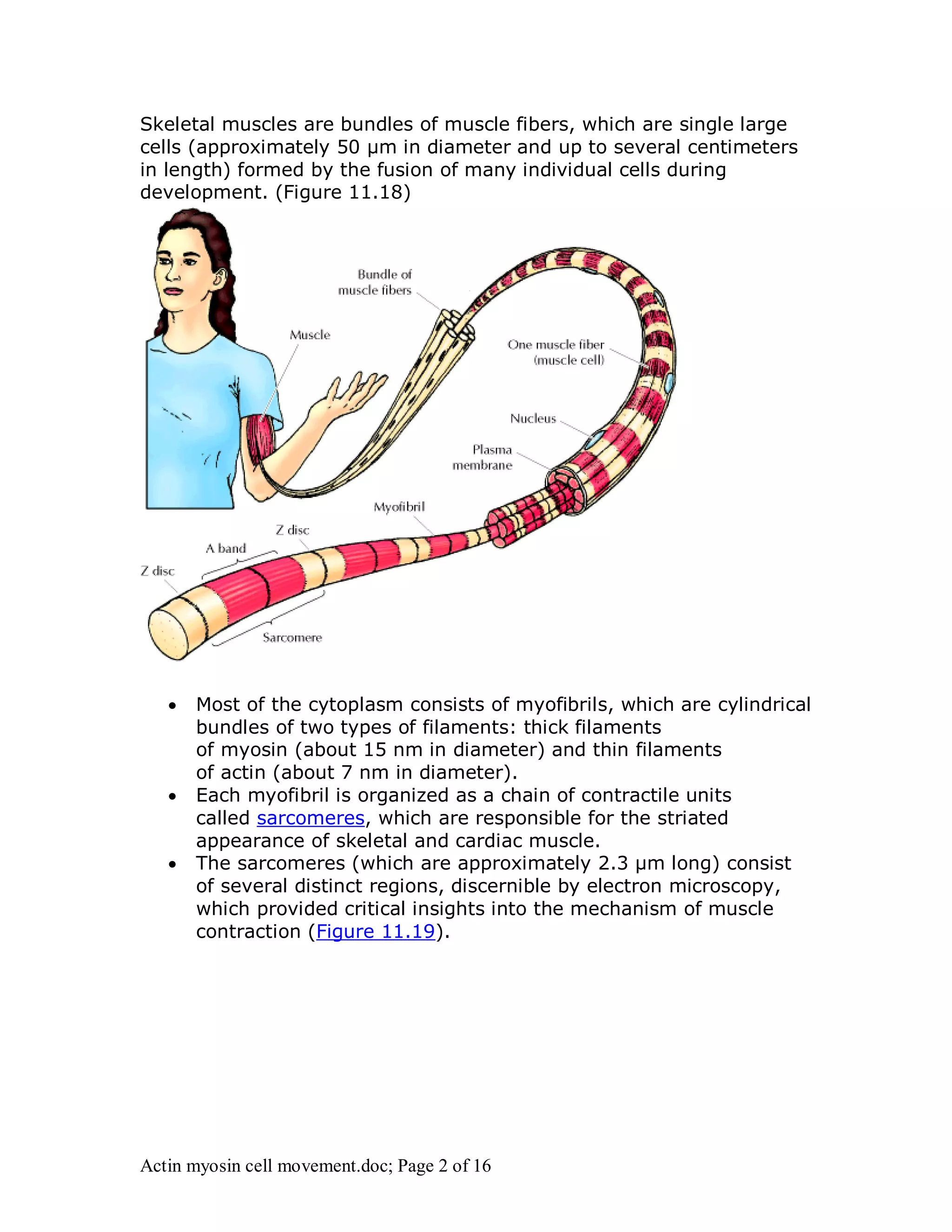




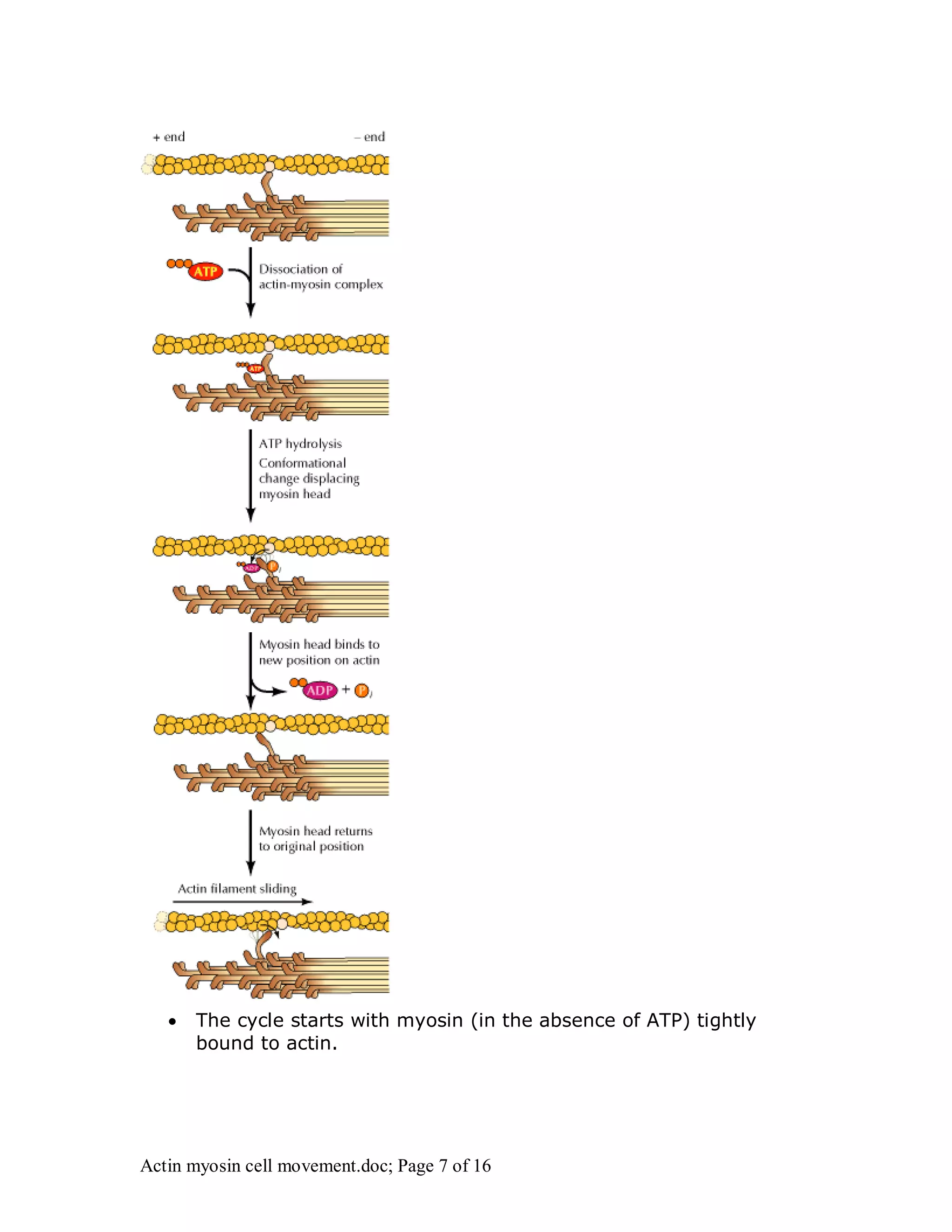

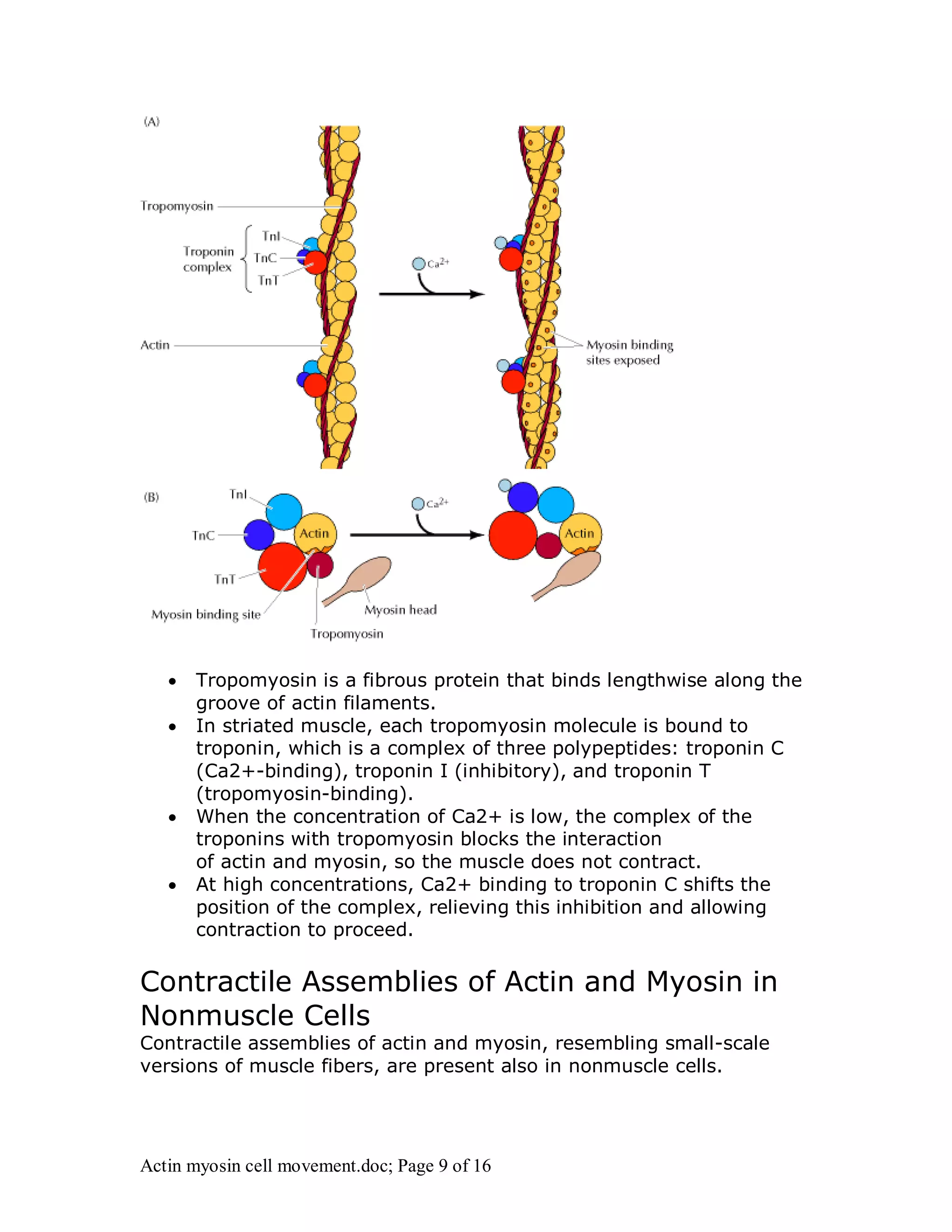

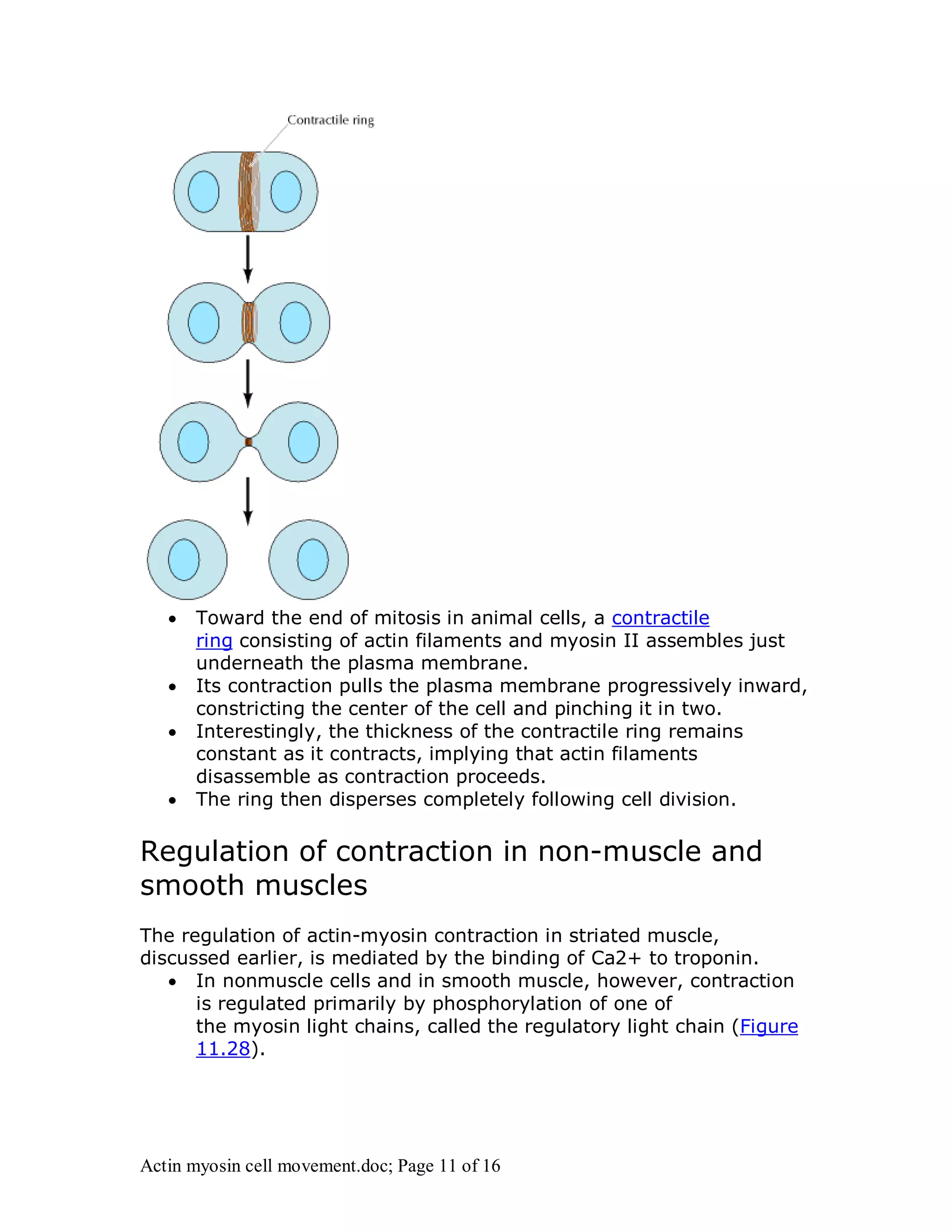
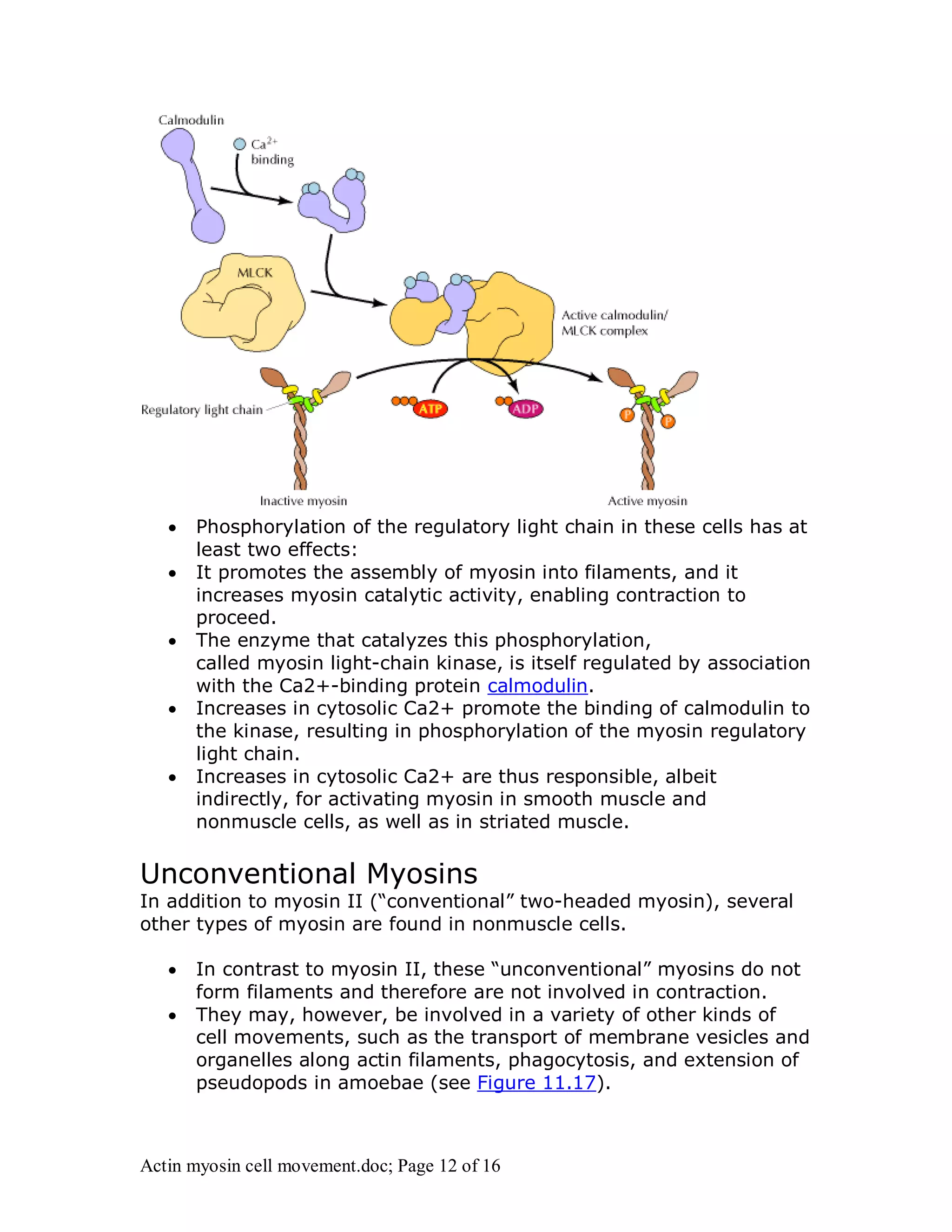





Actin and myosin interact to facilitate various cell movements, most notably muscle contraction, which is driven by the binding of myosin to actin filaments. The sliding filament model describes how muscle contraction occurs through the sliding of actin past myosin filaments within sarcomeres, ultimately resulting in the shortening of muscles. Additionally, actin-myosin interactions are vital for processes in nonmuscle cells, including cytokinesis and cell crawling.






































































































