Downloaded 242 times




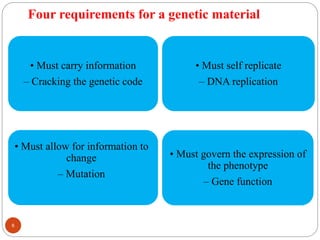
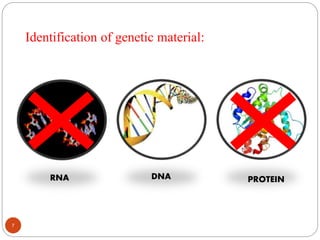

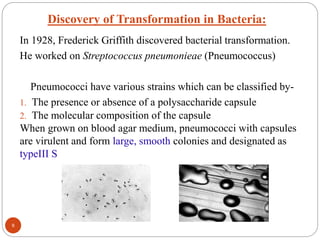
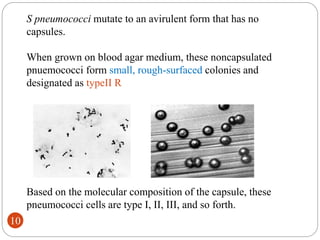
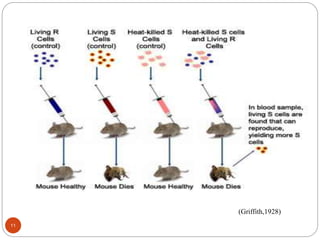


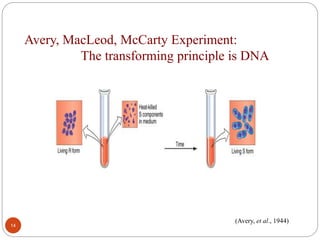
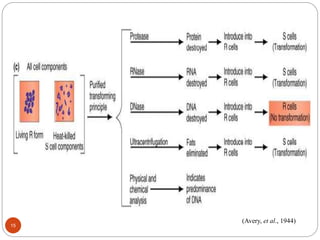

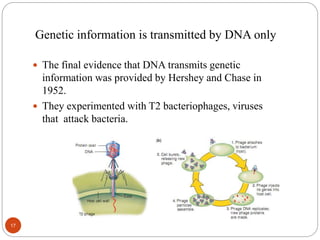
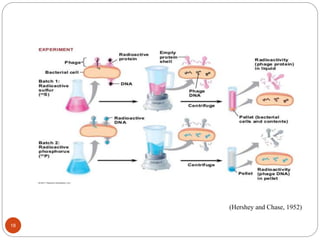
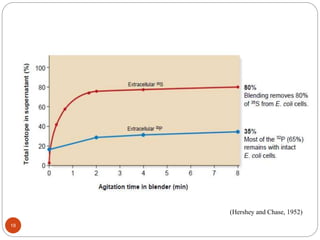

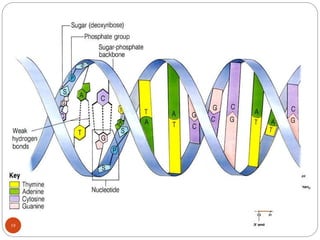
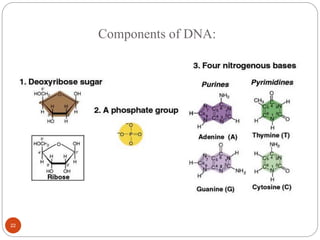
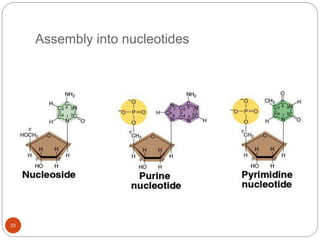
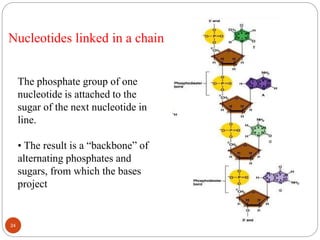
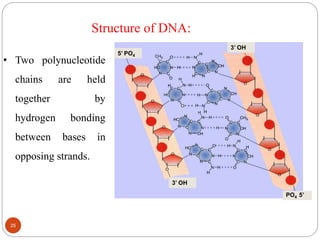
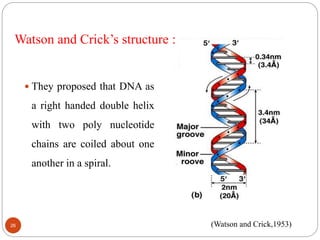


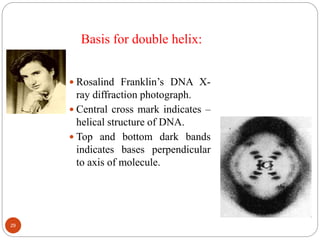
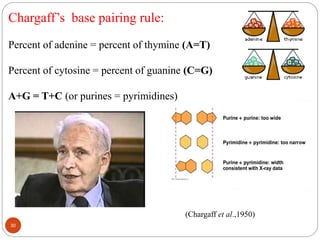


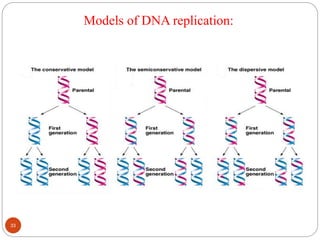
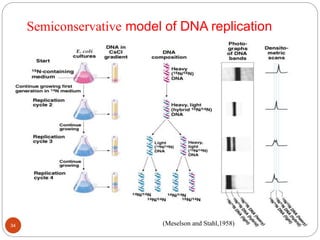

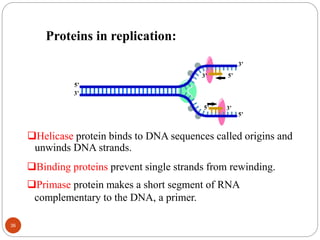
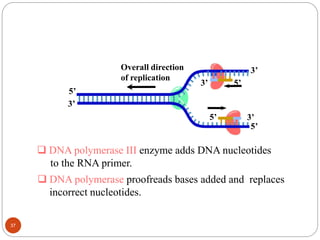
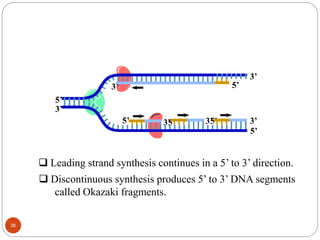
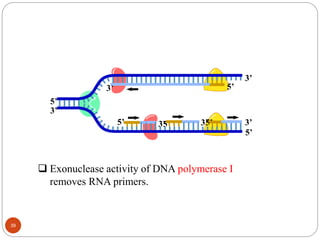
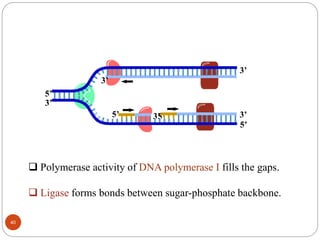
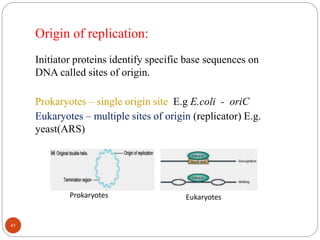

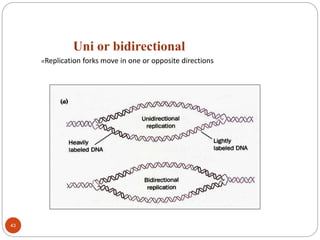
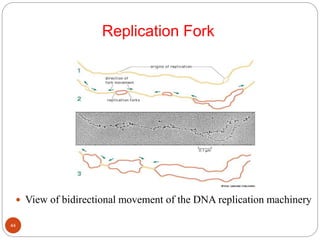
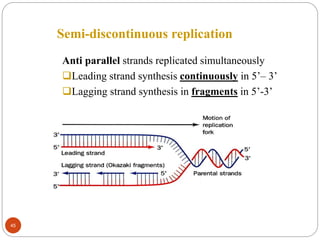
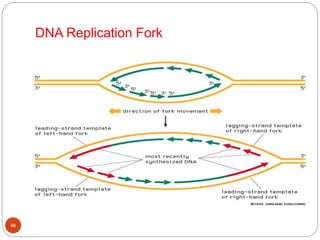
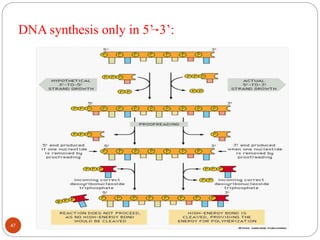

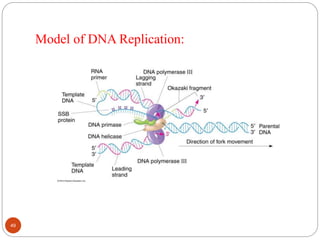
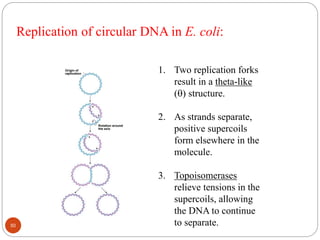
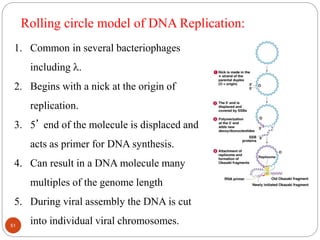


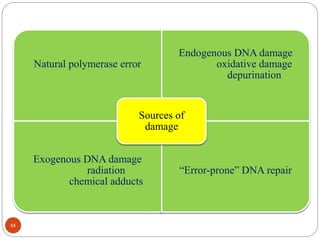


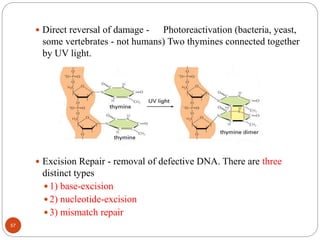
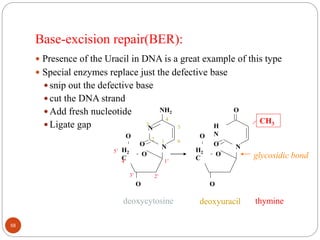
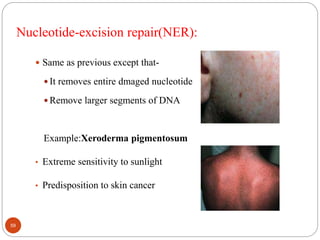



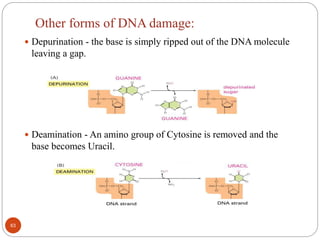




The document provides information on DNA, including its identification as the genetic material, its structure and replication. Some key points: - Experiments in the 1920s-1950s identified DNA as the genetic material, including Griffith's work on bacterial transformation and the experiments of Avery, MacLeod and McCarty and Hershey and Chase. - DNA is made up of nucleotides containing phosphate groups, sugars and nitrogenous bases. Watson and Crick discovered its double helix structure in 1953, with two anti-parallel strands held together by hydrogen bonds between complementary bases. - DNA replication is semi-conservative and involves unwinding of the DNA strands, synthesis of new complementary strands, and production of identical double























































































![Polymer [ बहुलक ] Chemistry Notes PDF - Irfanullah Mehar - JJ Sir Chemistry.pdf](https://cdn.slidesharecdn.com/ss_thumbnails/polymerchemistrynotespdf-irfanullahmehar-jjsirchemistry-260210172118-3f9b37f7-thumbnail.jpg?width=640&height=640&fit=bounds)











