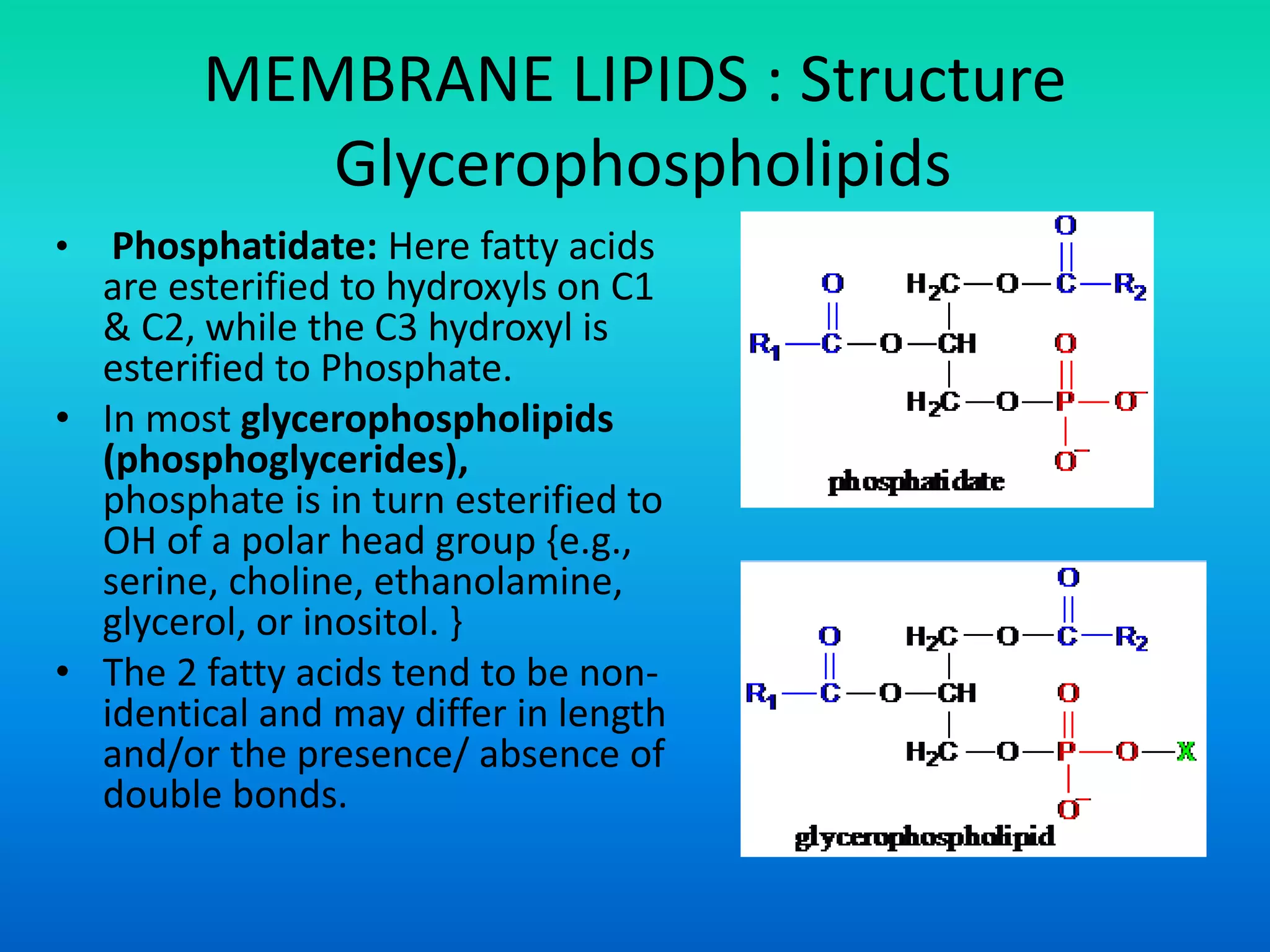
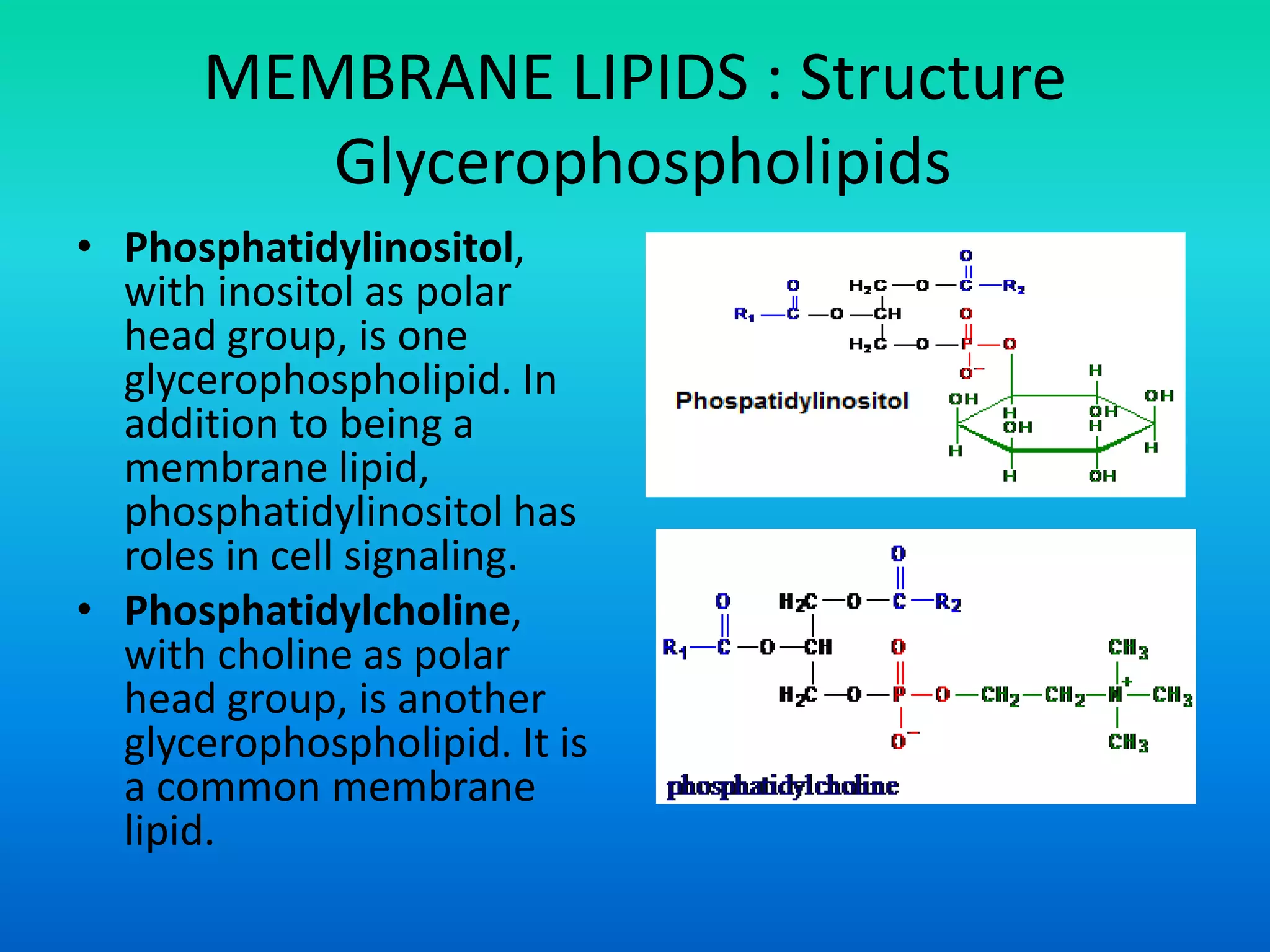
This document discusses the structure and functions of membrane lipids. It describes the main types of membrane lipids - glycerophospholipids, sphingolipids, glycosphingolipids, and cholesterol. It explains the structures of these lipids, including their polar head groups and non-polar hydrocarbon tails which allow them to form lipid bilayers. The document also summarizes how membrane lipids contribute to membrane fluidity and permeability, noting how cholesterol and unsaturated fatty acids influence fluidity and prevent crystallization.