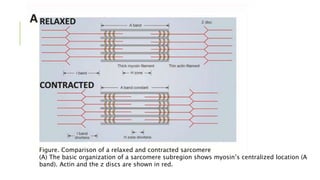
- Muscle contraction occurs via the sliding filament theory where actin filaments slide past myosin filaments, causing sarcomeres and muscles to shorten.
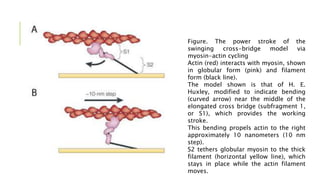
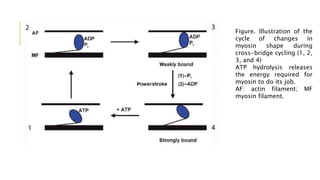
- Sarcomeres contain regularly arranged thick and thin filaments that generate striations visible under a microscope. Contraction is driven by myosin cross-bridges binding to actin and generating a power stroke via ATP hydrolysis.
- Calcium binds to troponin and causes it to shift tropomyosin, unblocking the myosin binding sites on actin and allowing cross-bridge cycling and contraction to occur.
























































![2022 bio20-8a - MuscularSystemreview [ST].pdf](https://cdn.slidesharecdn.com/ss_thumbnails/2022bio20-8a-muscularsystemst-250521201551-86d3a57c-thumbnail.jpg?width=640&height=640&fit=bounds)









































