Downloaded 181 times
















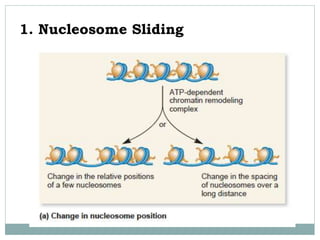

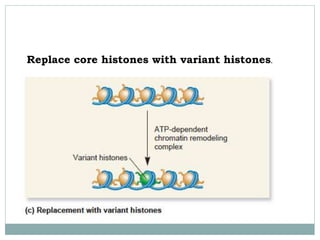
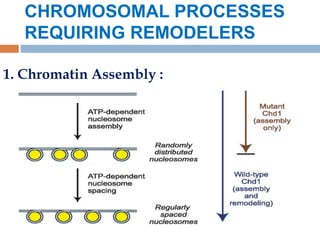

















1. Chromatin remodeling is the process by which chromatin structure is dynamically modified to allow access of DNA for processes like transcription. 2. There are two main types of chromatin remodeling - covalent histone modification and ATP-dependent chromatin remodeling complexes. 3. ATP-dependent complexes use energy from ATP hydrolysis to move, eject, or restructure nucleosomes, allowing access to DNA. 4. Examples of chromatin remodeling complexes include SWI/SNF, ISWI, CHD, and INO80 families, which have different activities like nucleosome sliding or histone variant exchange.










































![New_MY_Resume_2015[1]](https://cdn.slidesharecdn.com/ss_thumbnails/b964ae14-03dd-4dbd-87b3-ba0438e9b3d0-160426042900-thumbnail.jpg?width=640&height=640&fit=bounds)



















