Downloaded 228 times





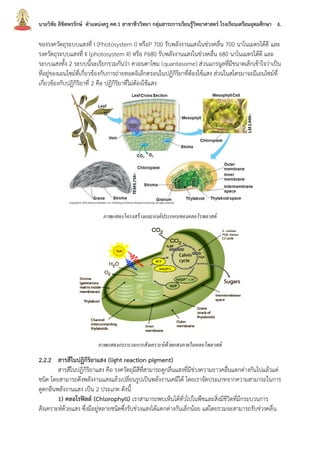

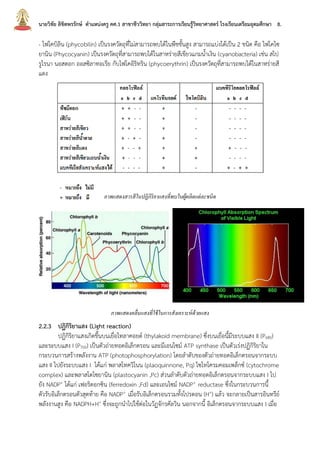

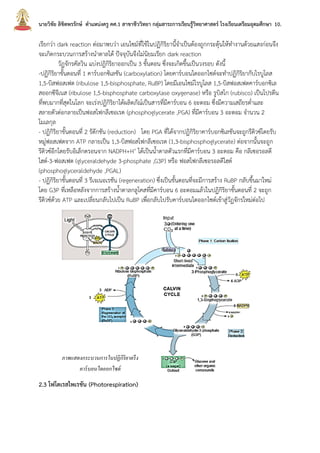

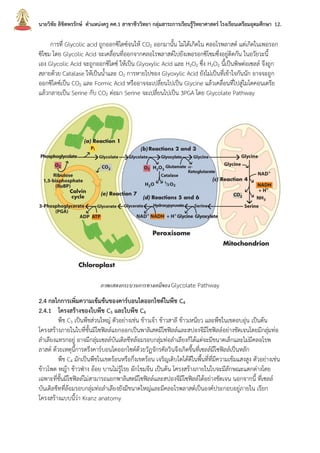
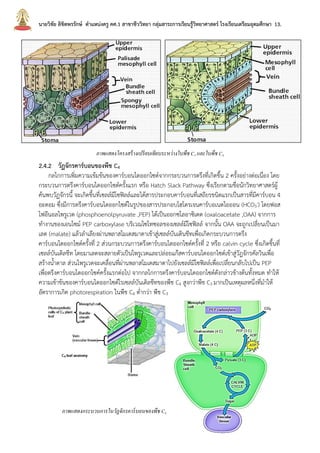

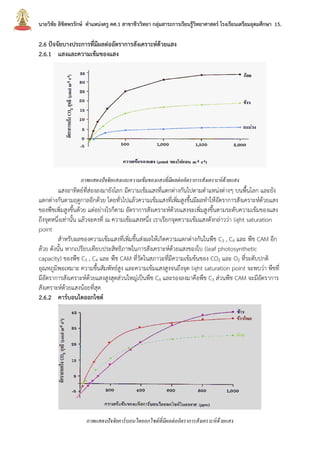
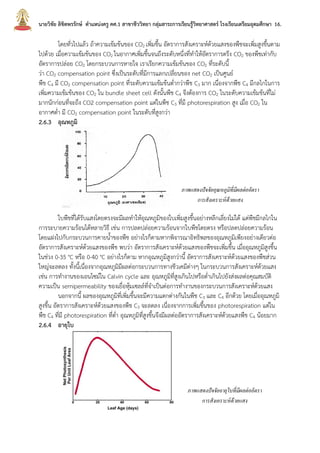




บทที่2กระบวนการสังเคราะห์ด้วยแสง นายวิชัย ลิขิตพรรักษ์ ตำแหน่งครู คศ.1 เอกวิชาชีววิทยา ประวัติการศึกษา : พ.ศ. 2549 วิทยาศาสตรบัณฑิต (เกีรยตินิยมอันดับ 2) สาขาชีววิทยา คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล พ.ศ. 2551 ศึกษาศาสตรบัณฑิต สาขาวิชาศึกษาศาสตร์ เอกเทคโนโลยีและสื่อสารการศึกษา มหาวิทยาลัยสุโขทัยธรรมาธิราช พ.ศ. 2552 ประกาศนียบัตรบัณฑิตวิชาชีพครู คณะครุศาสตร์ มหาวิทยาลัยราชภัฏสวนดุสิต พ.ศ. 2555 สาธารณสุขศาสตรบัณฑิต สาขาวิชาวิทยาศาสตร์สุขภาพ เอกสาธารณสุขศาสตร์ มหาวิทยาลัยสุโขทัยธรรมาธิราช พ.ศ. 2558 ศึกษาศาสตรมหาบัณฑิต สาขาวิชาการประเมินและการวิจัยทางการศึกษา เอกวิจัยทางการศึกษา คณะศึกษาศาสตร์ มหาวิทยาลัยรามคำแหง























































































