Downloaded 189 times


This document summarizes key aspects of voluntary motor control and the neural systems involved. It discusses how the basal ganglia, cerebellum, and motor cortex work together to plan and execute movements. The spinal cord and motor neurons are also covered, as well as mechanisms of muscle contraction and sensory feedback systems that provide input to modulate movement.
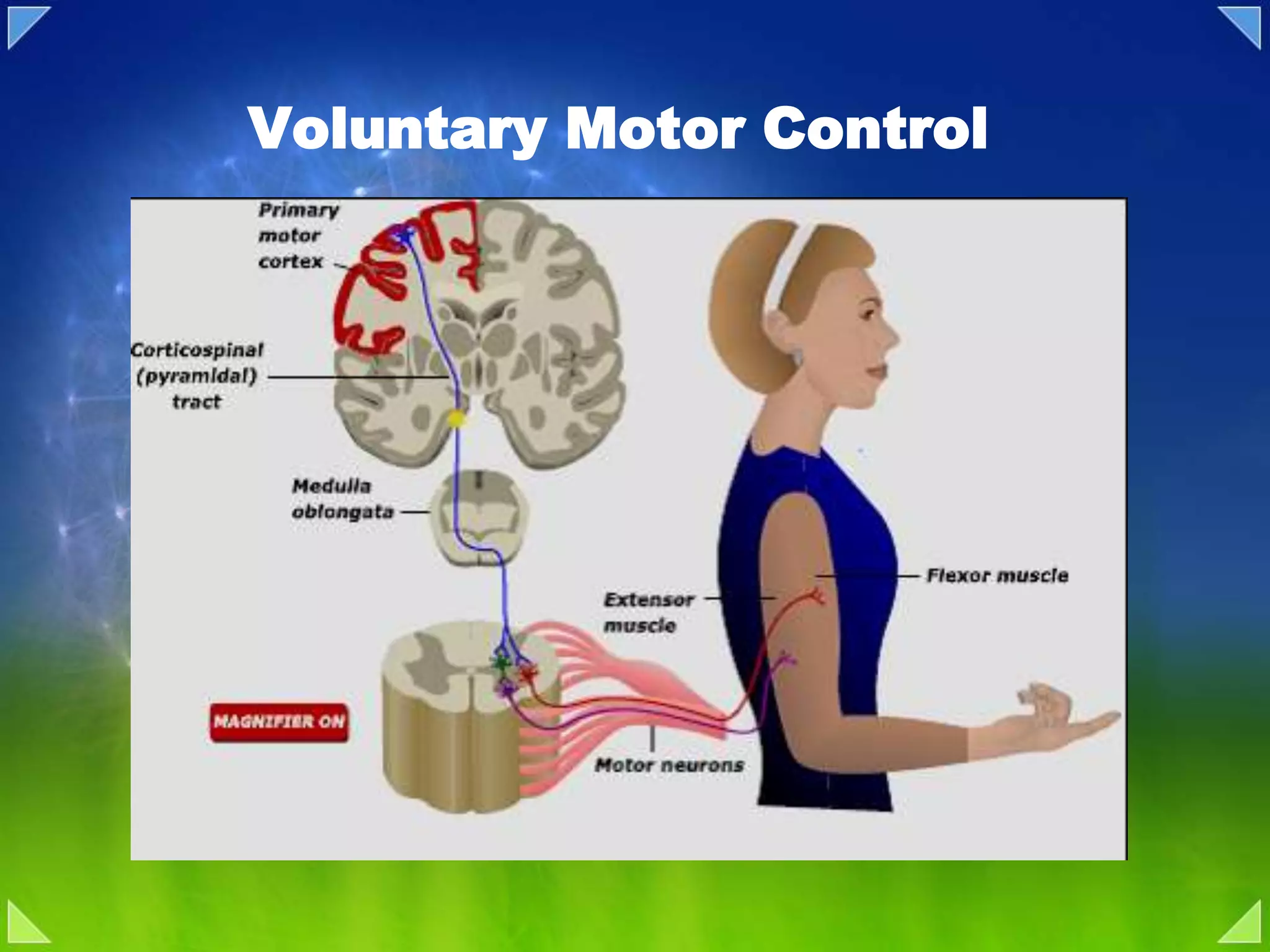
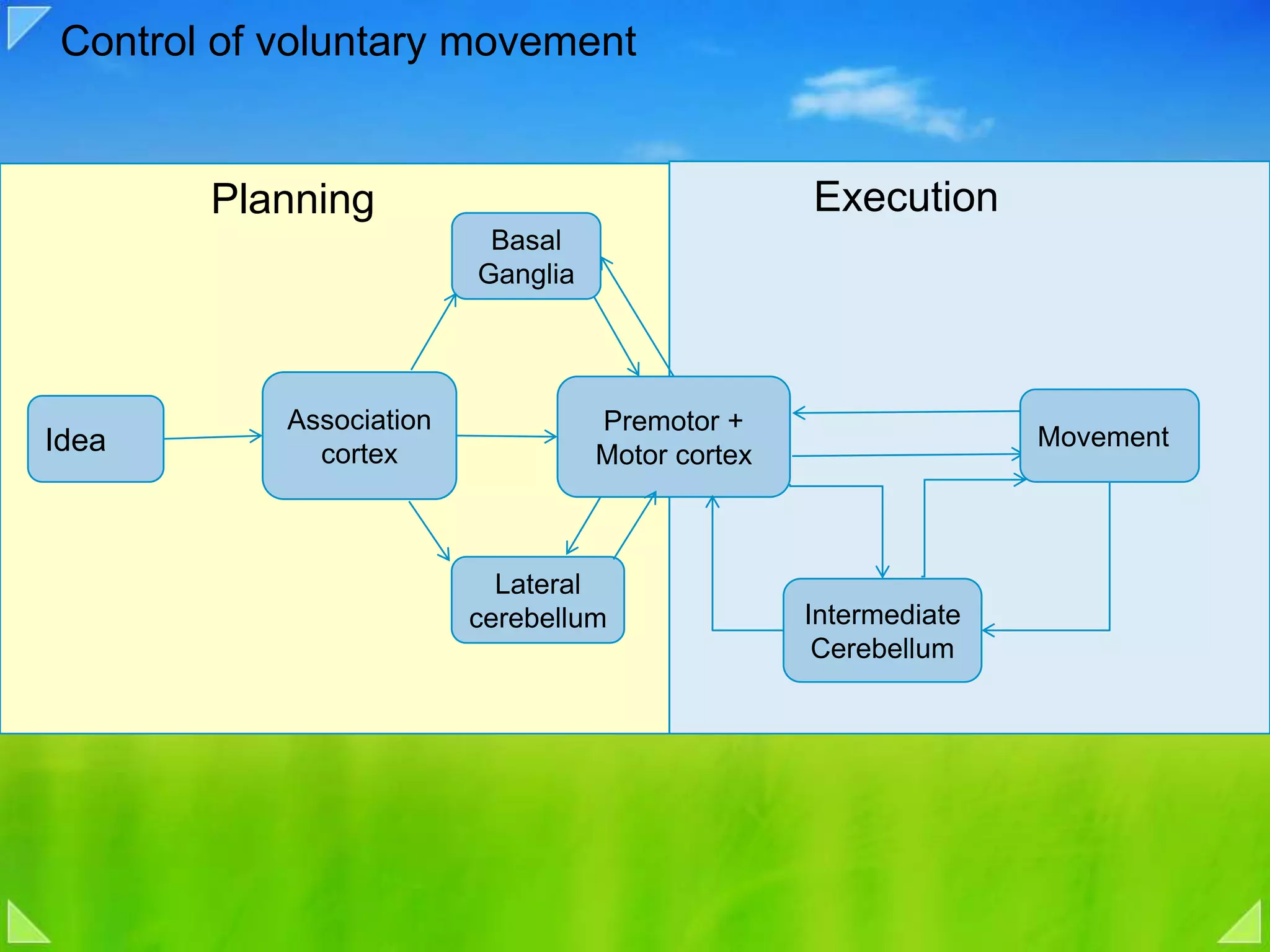
Introduction to voluntary motor control including execution, planning, and brain areas involved.
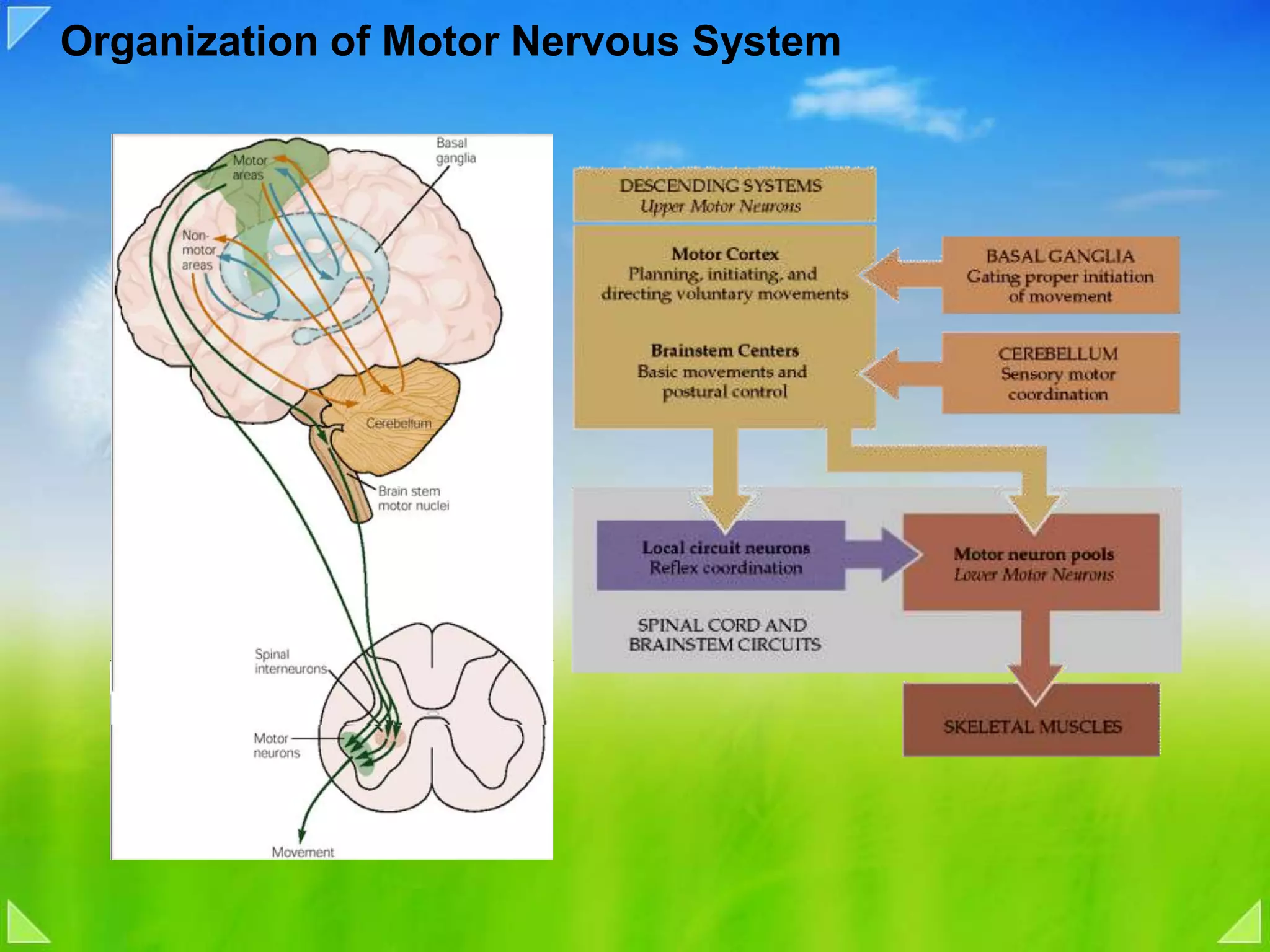
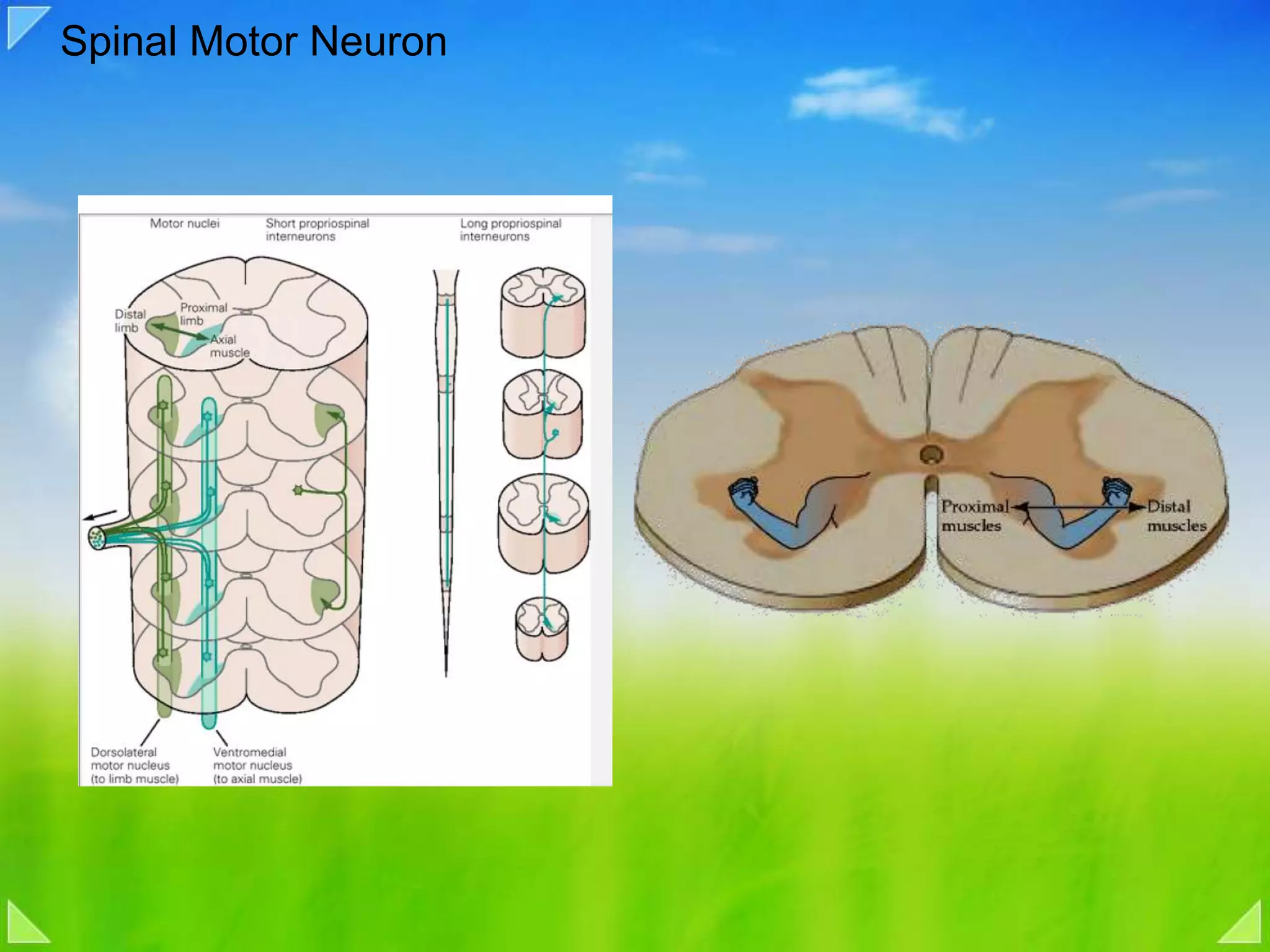
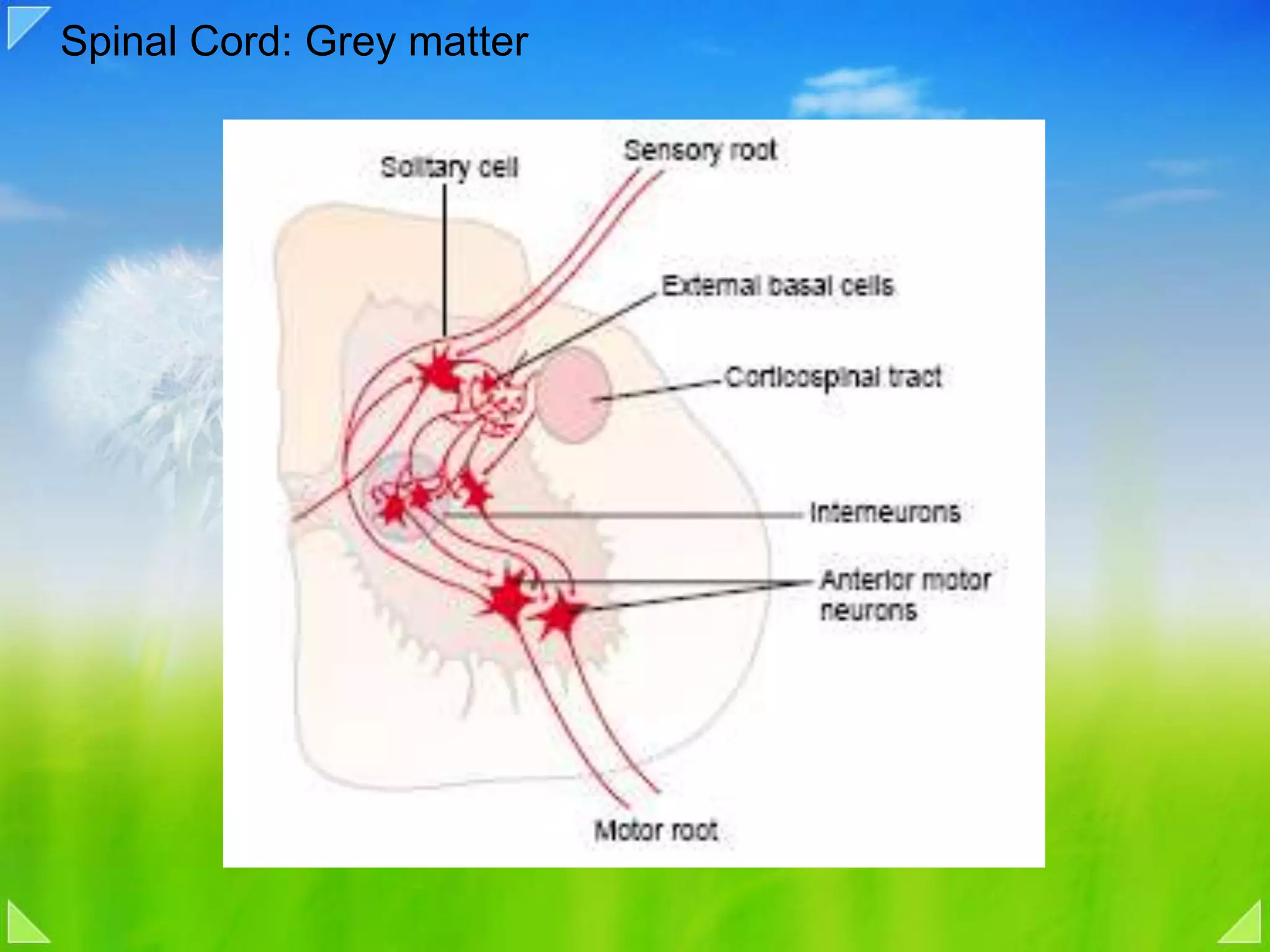
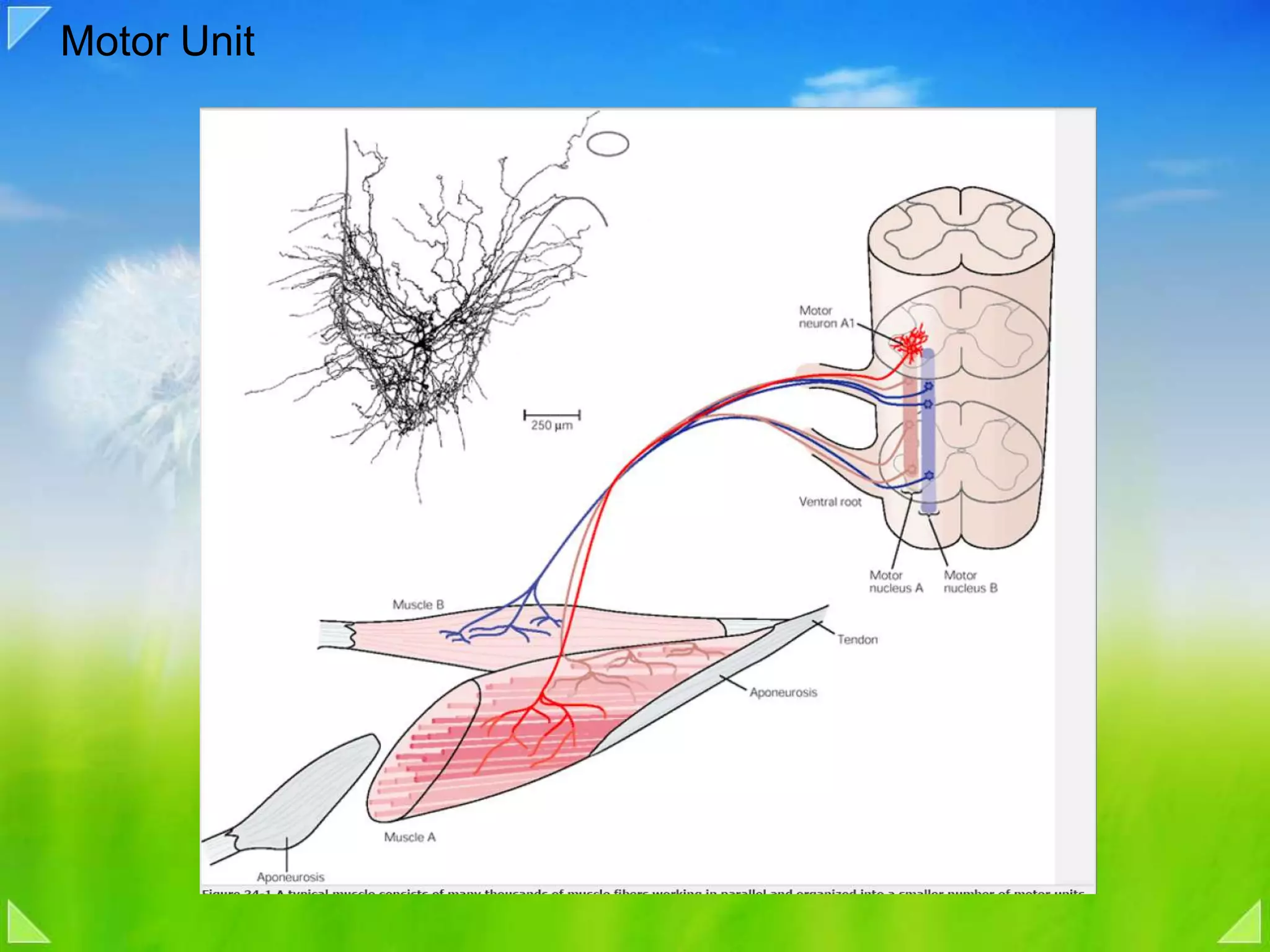
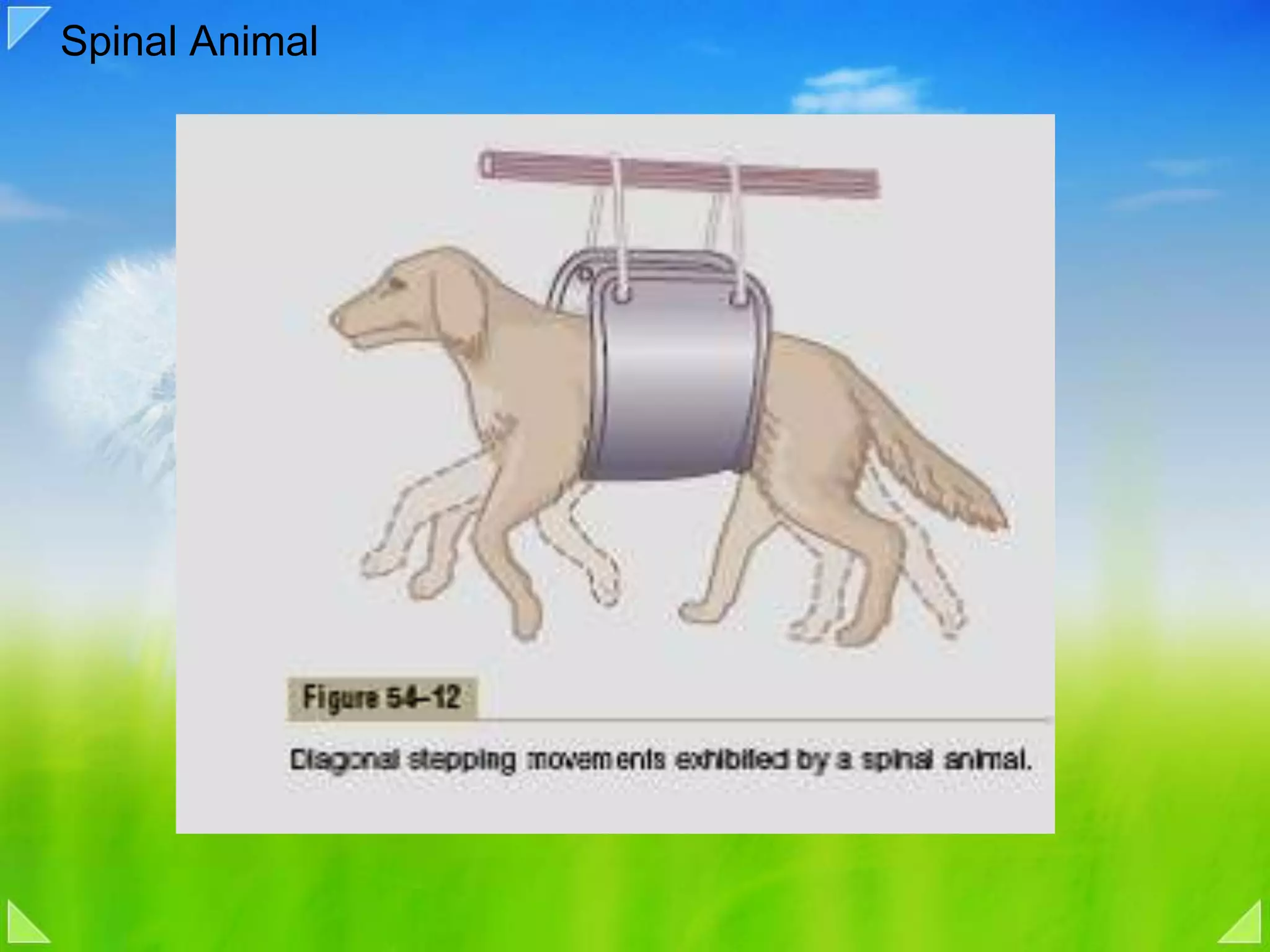
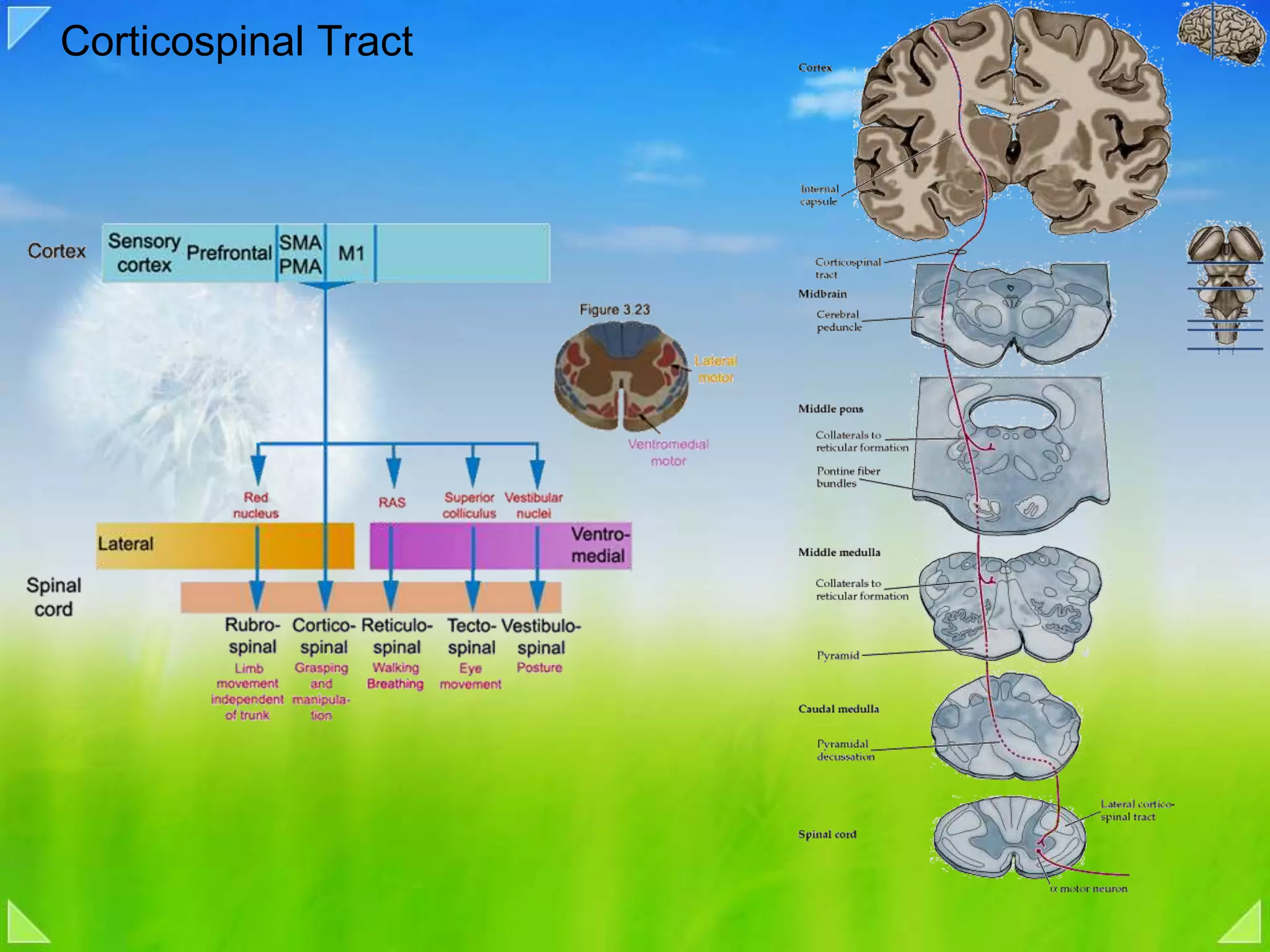
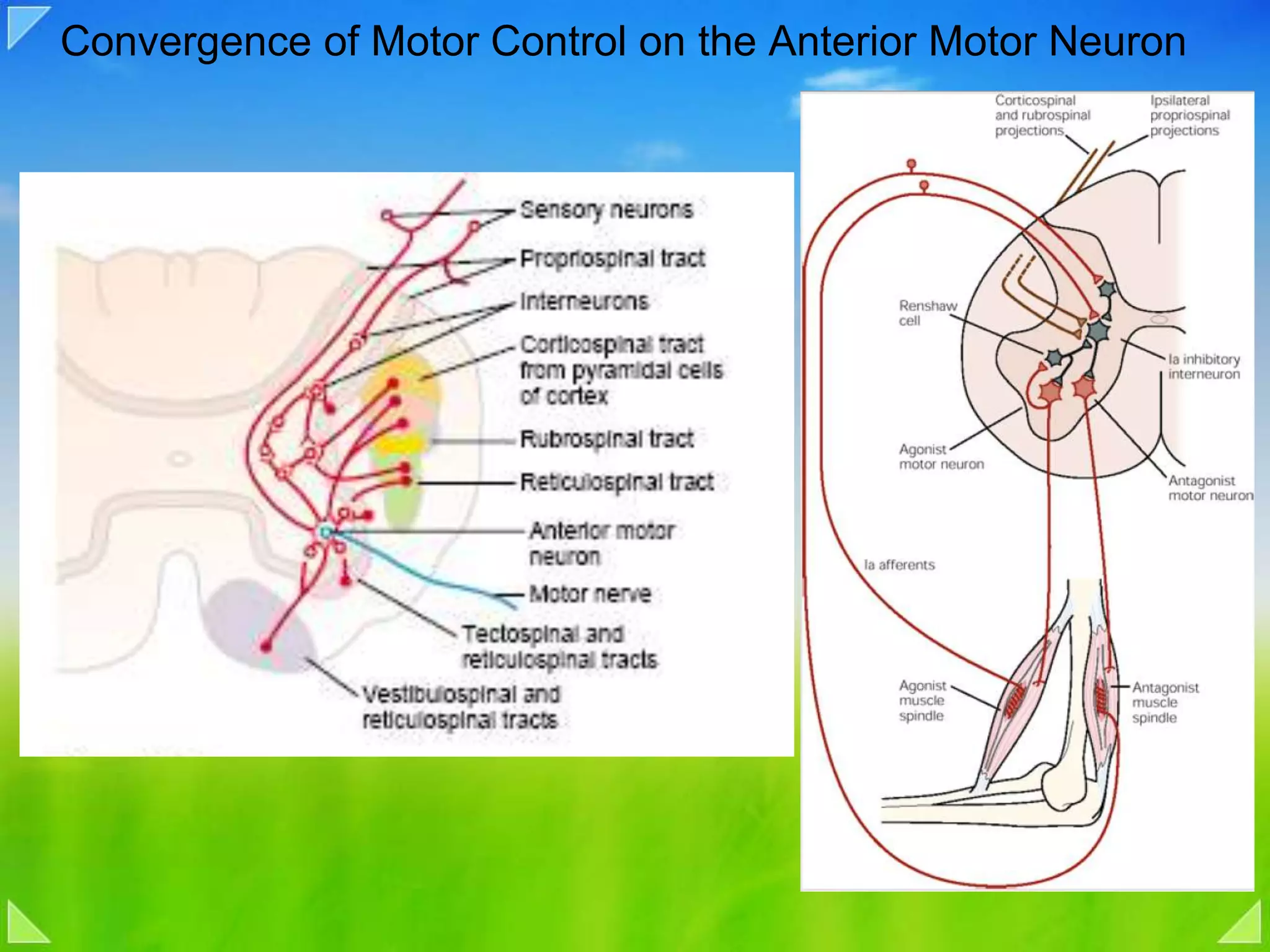
Organization of the motor nervous system, including spinal motor neurons, spinal cord structure, and motor units.
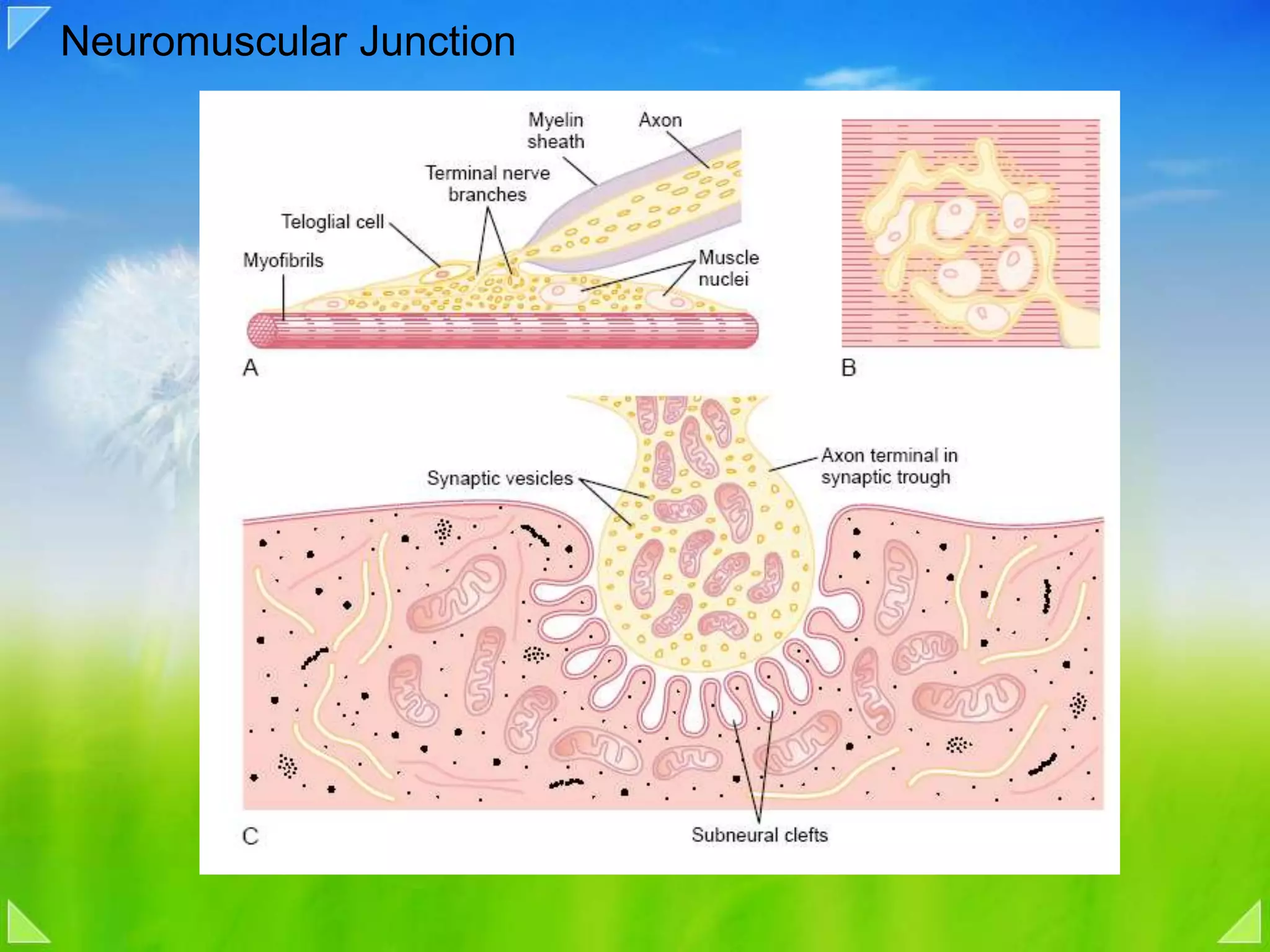
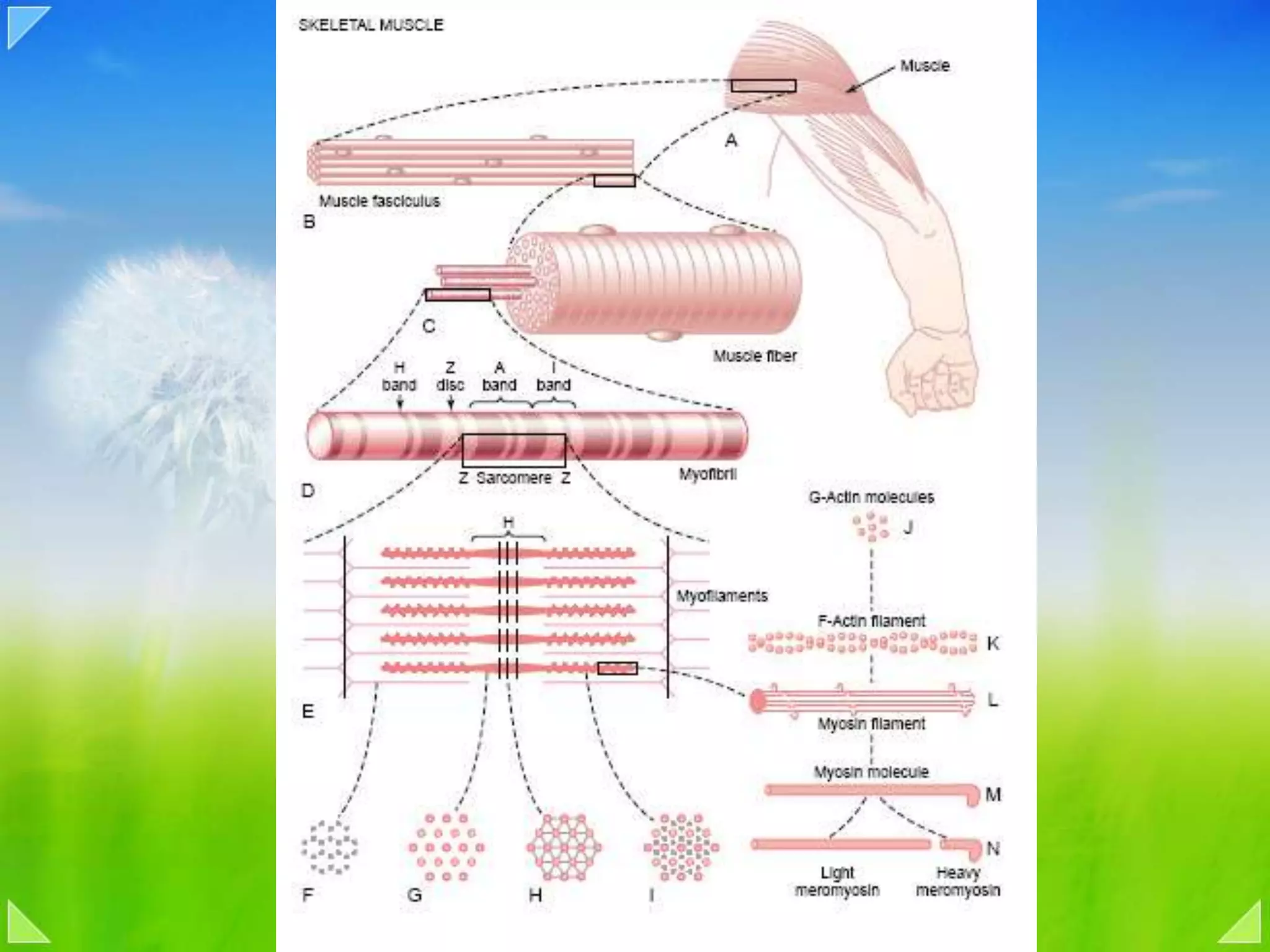
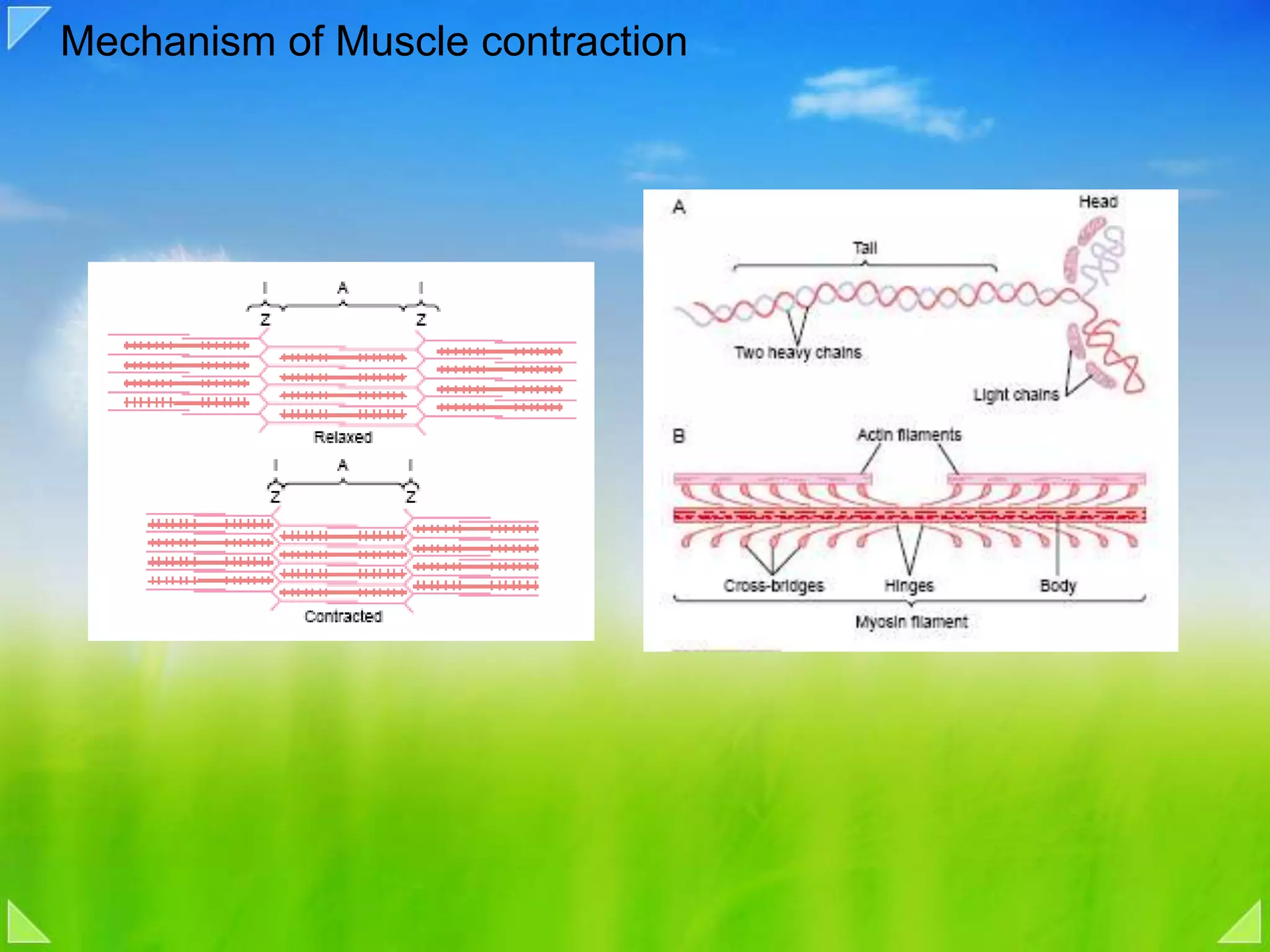
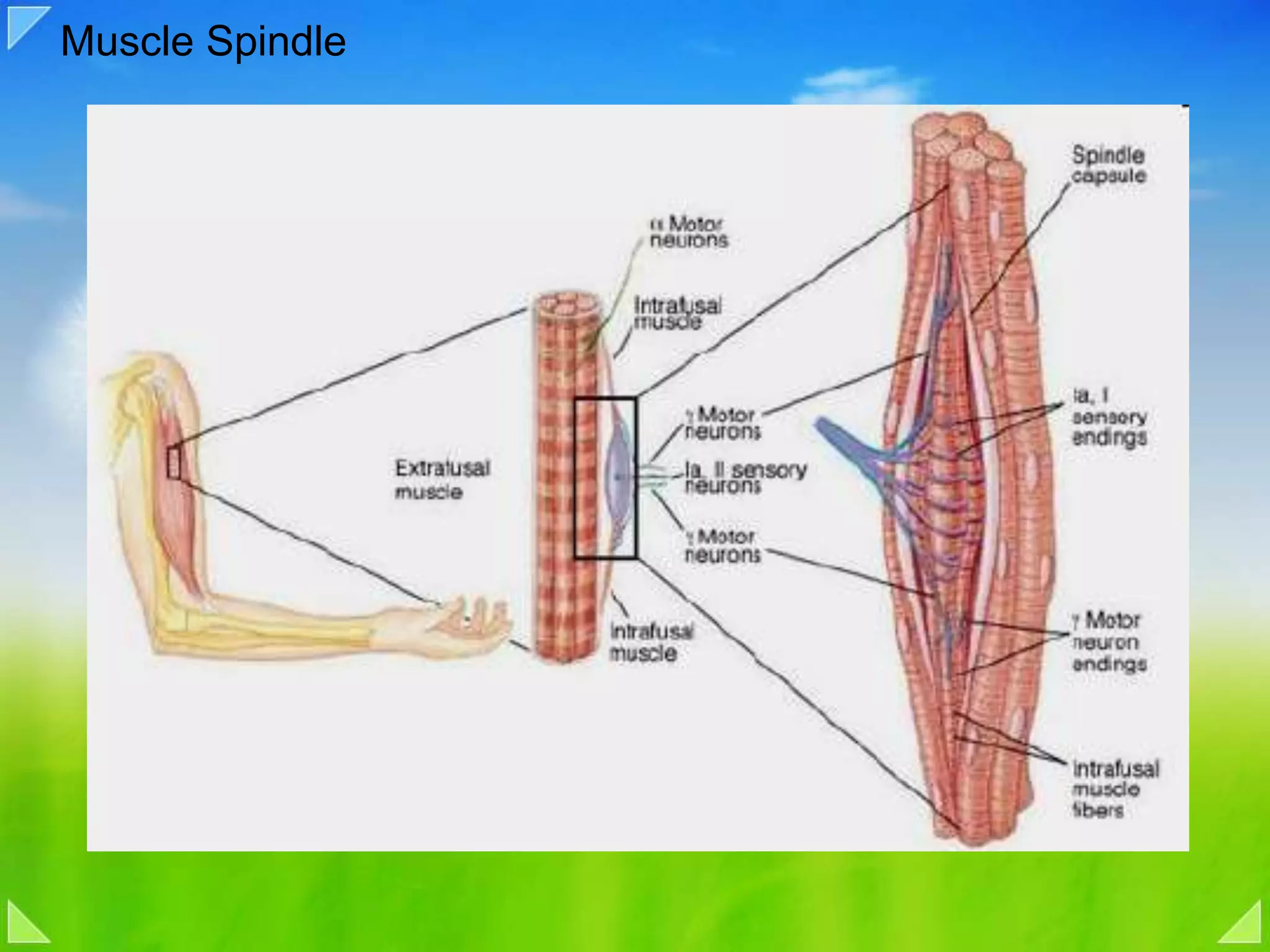
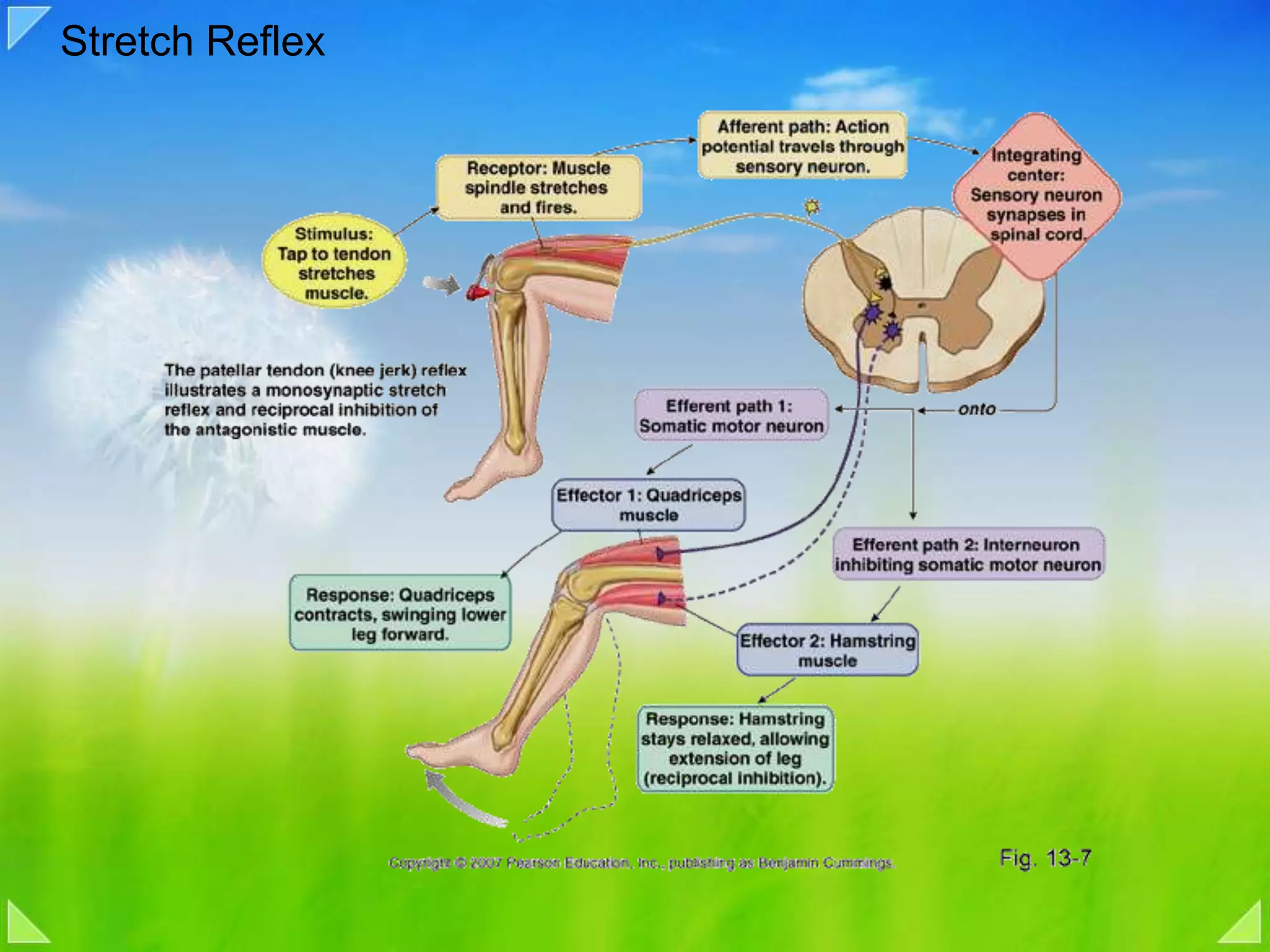
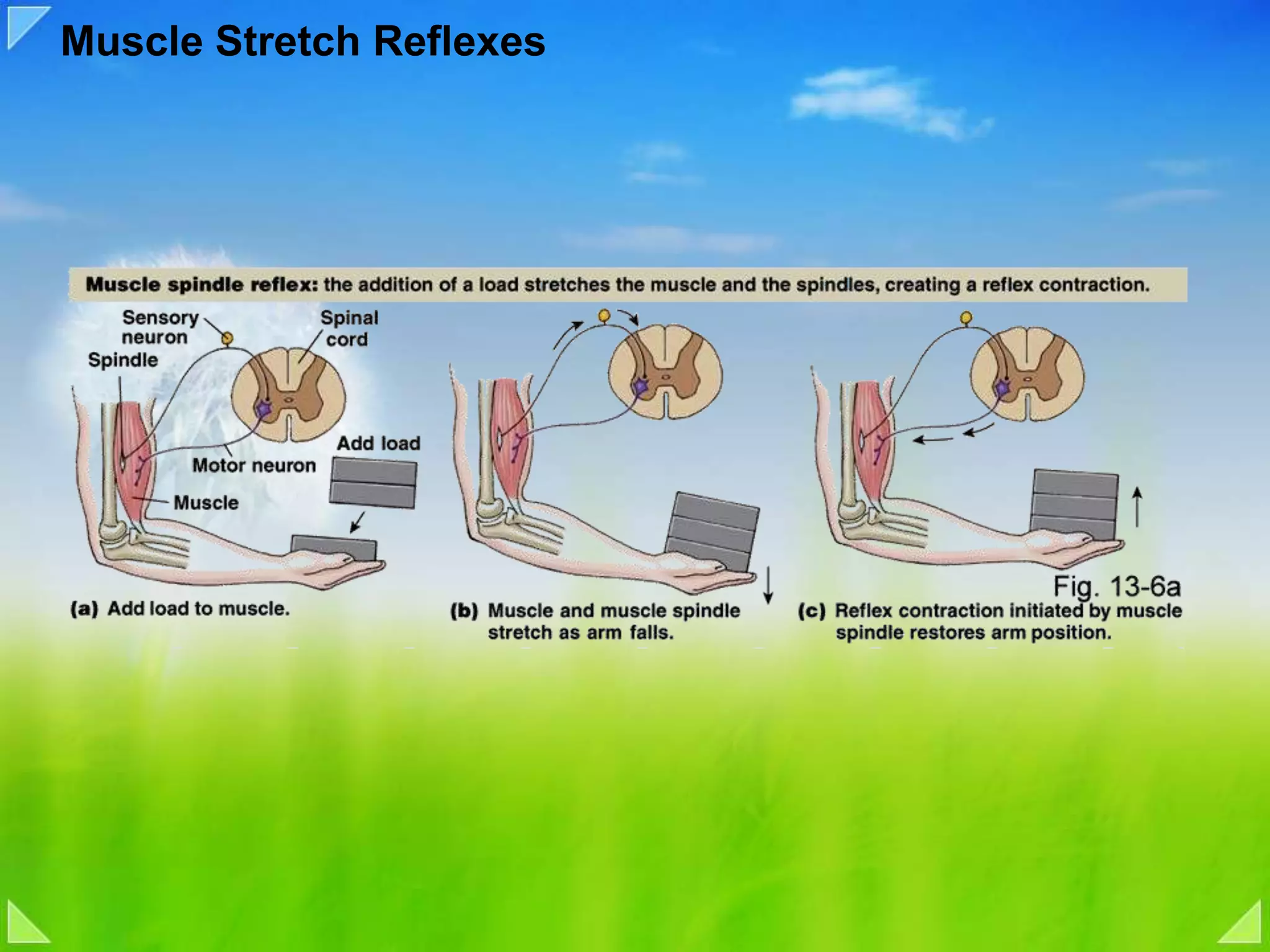
Details on the neuromuscular junction, muscle contraction mechanisms, and muscle stretch reflexes.
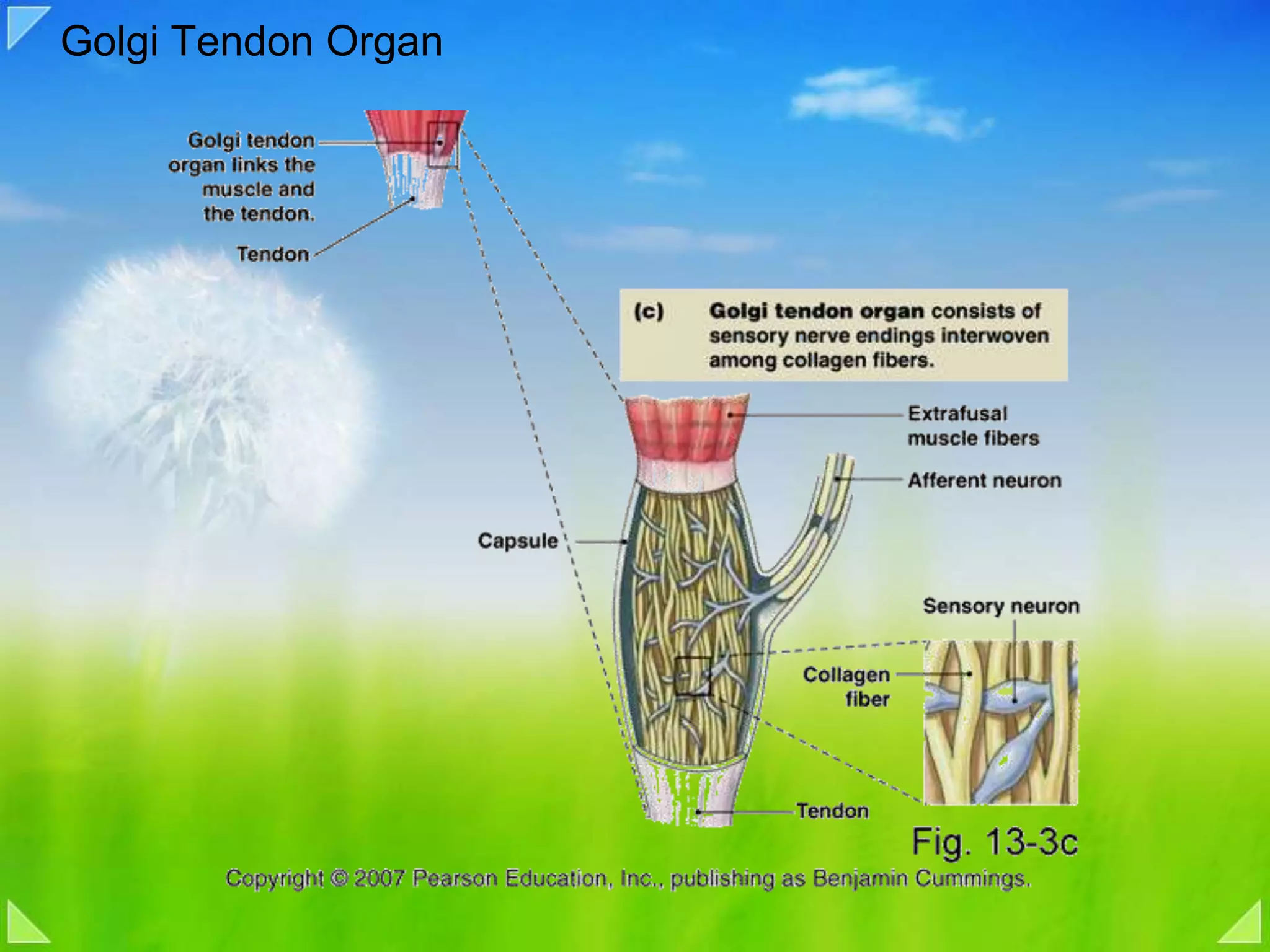
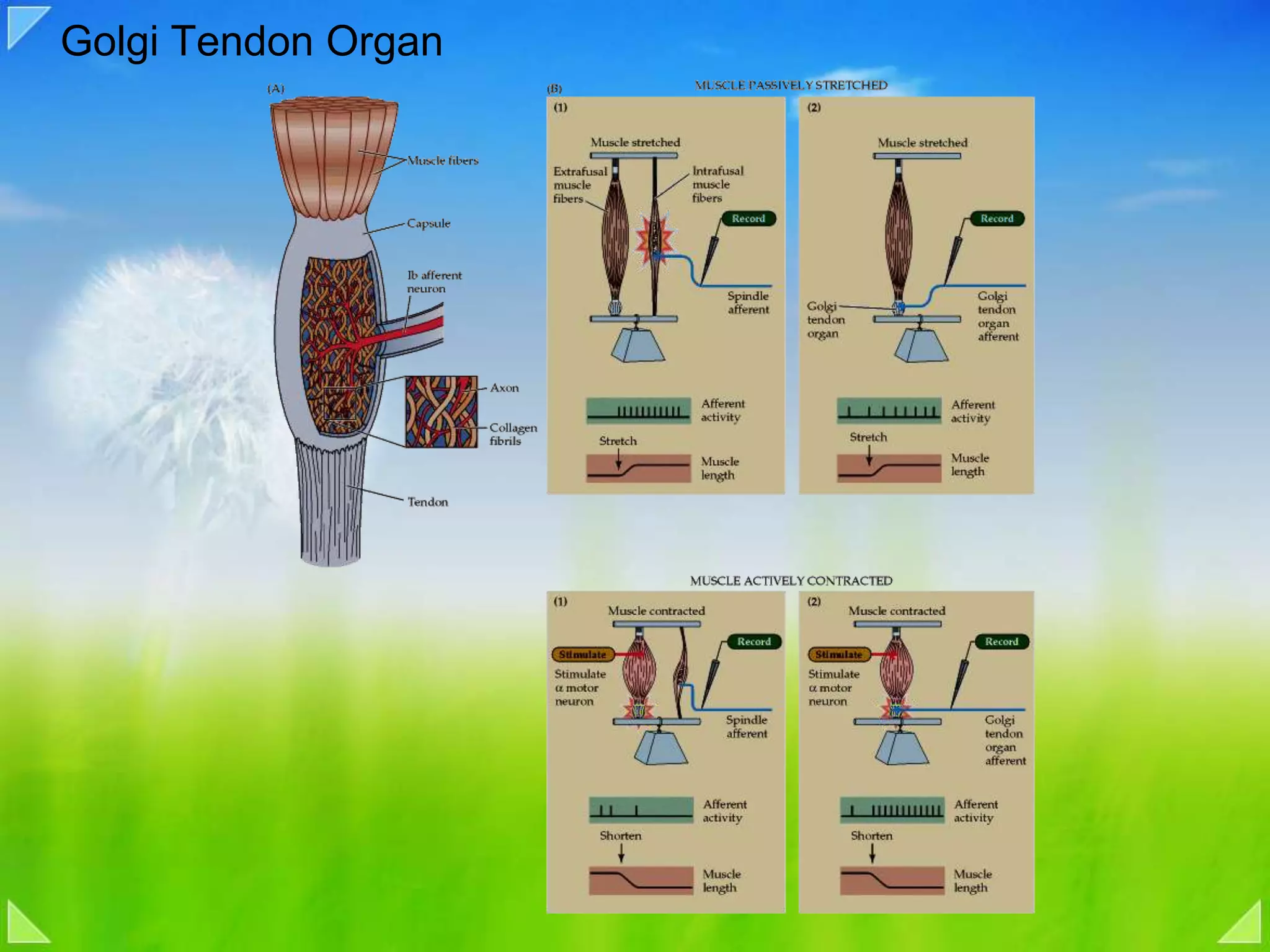
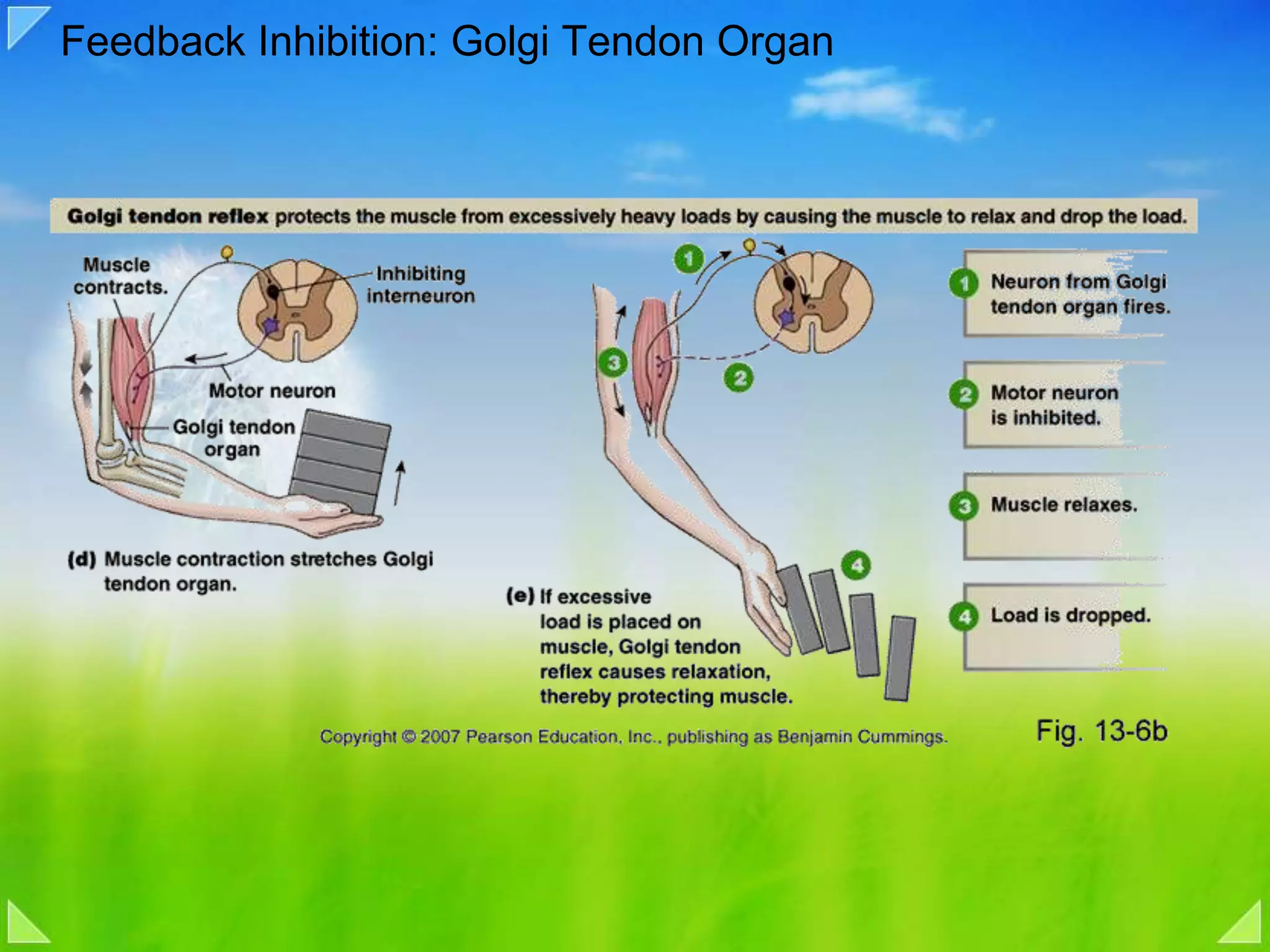
Functions of the Golgi tendon organ, feedback inhibition and its role in muscle regulation.
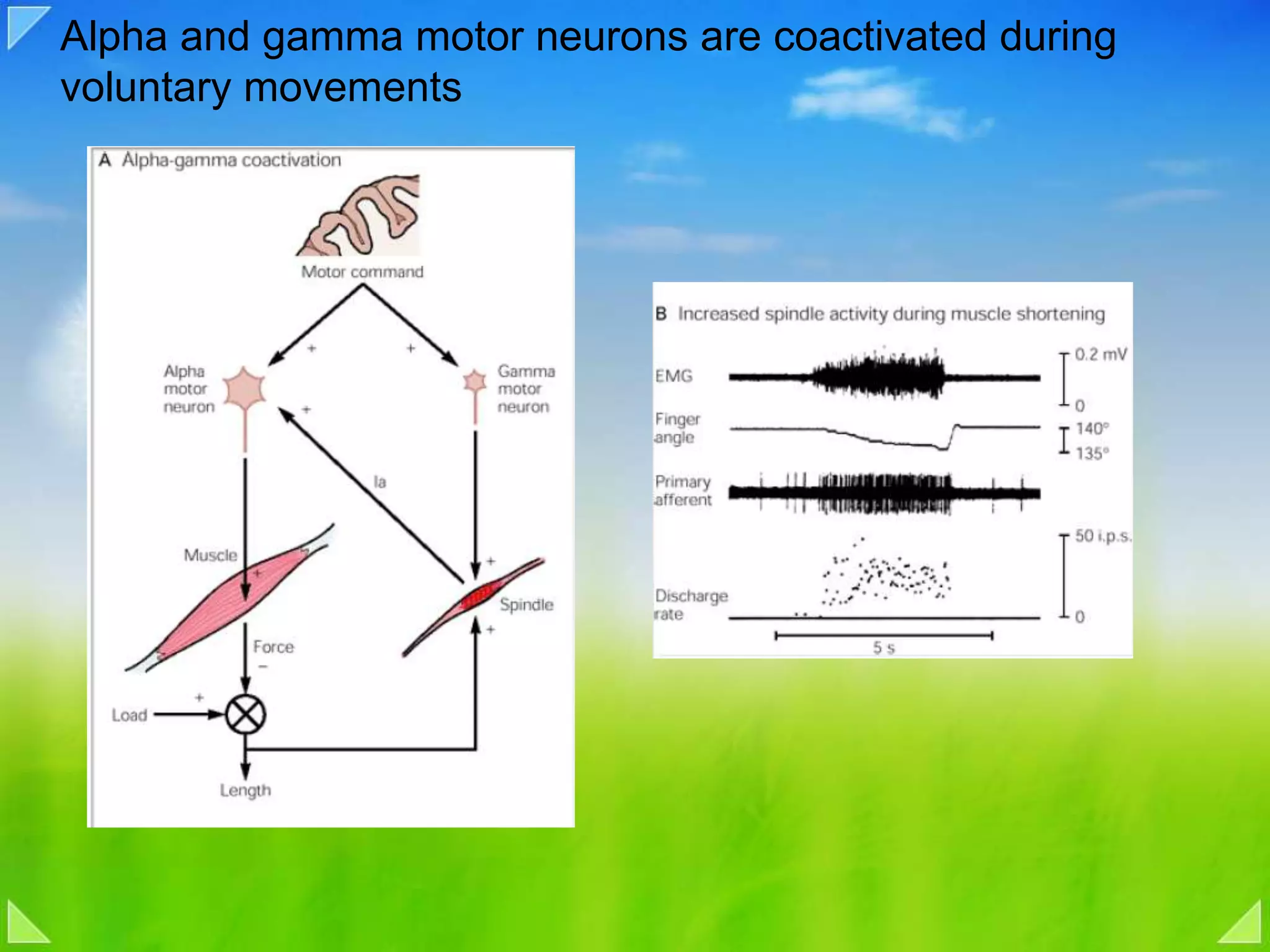
Coactivation of alpha and gamma motor neurons, and implications in voluntary movements.
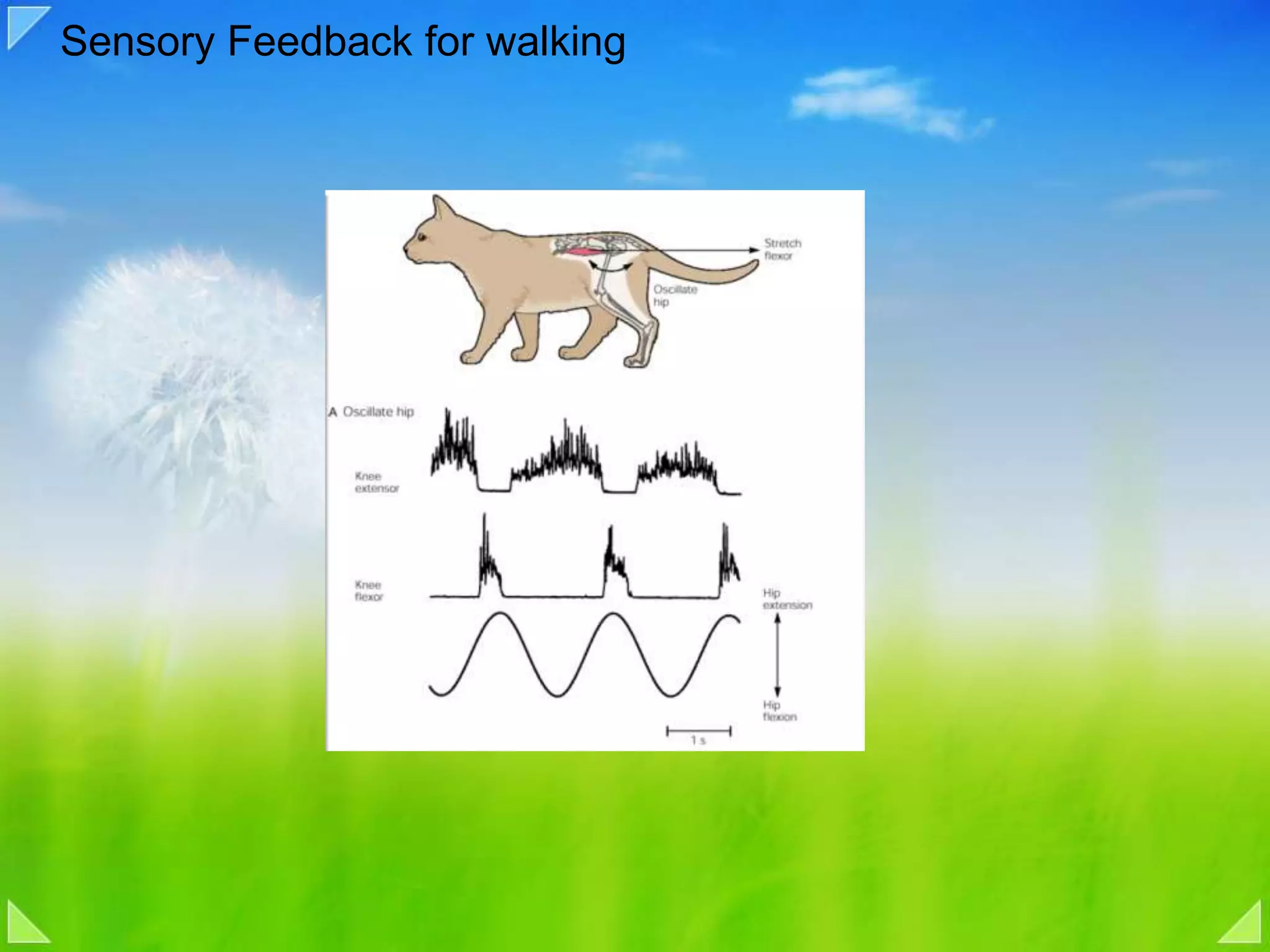
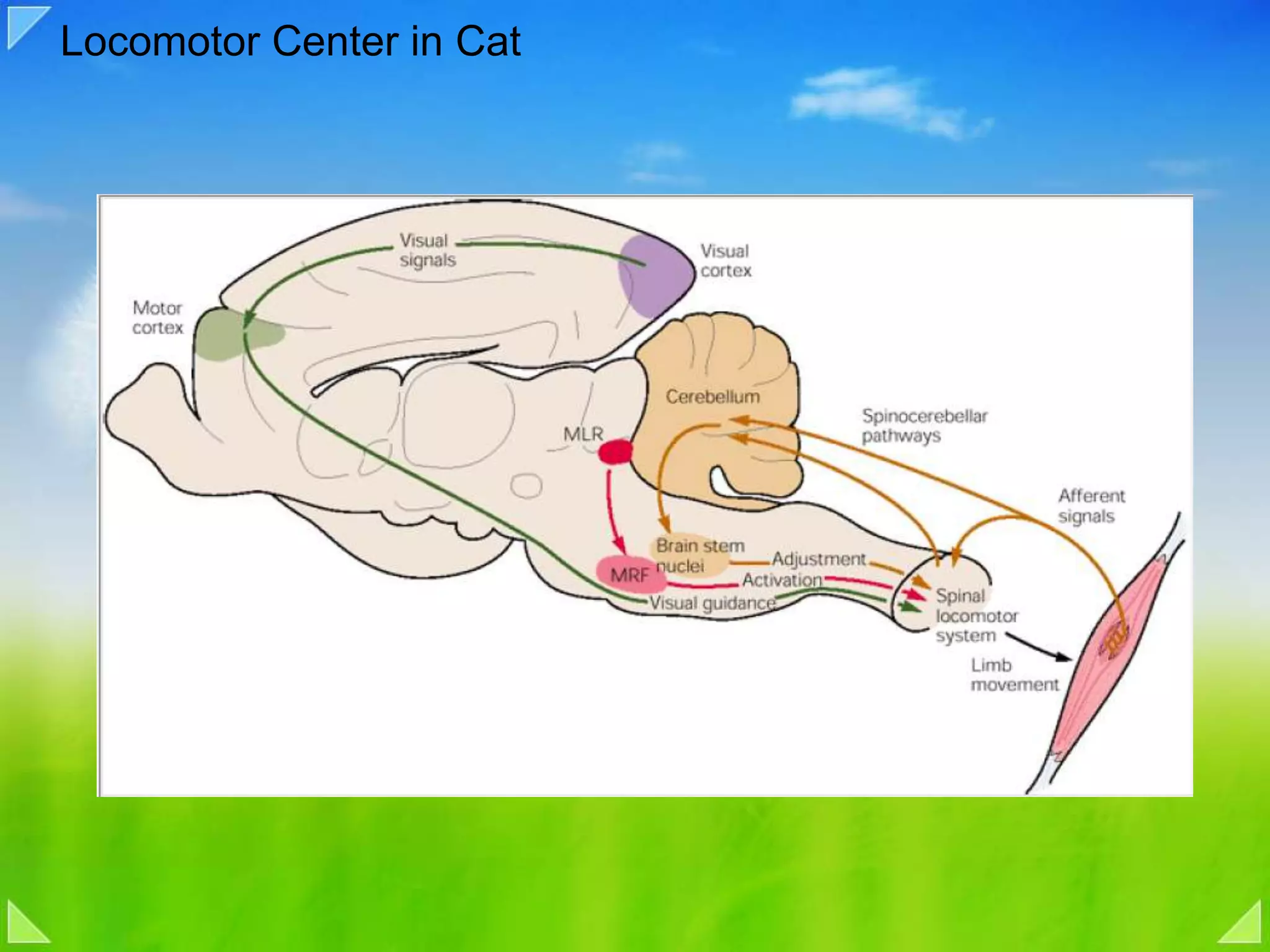
Role of sensory feedback and locomotor centers in walking, using cat studies.
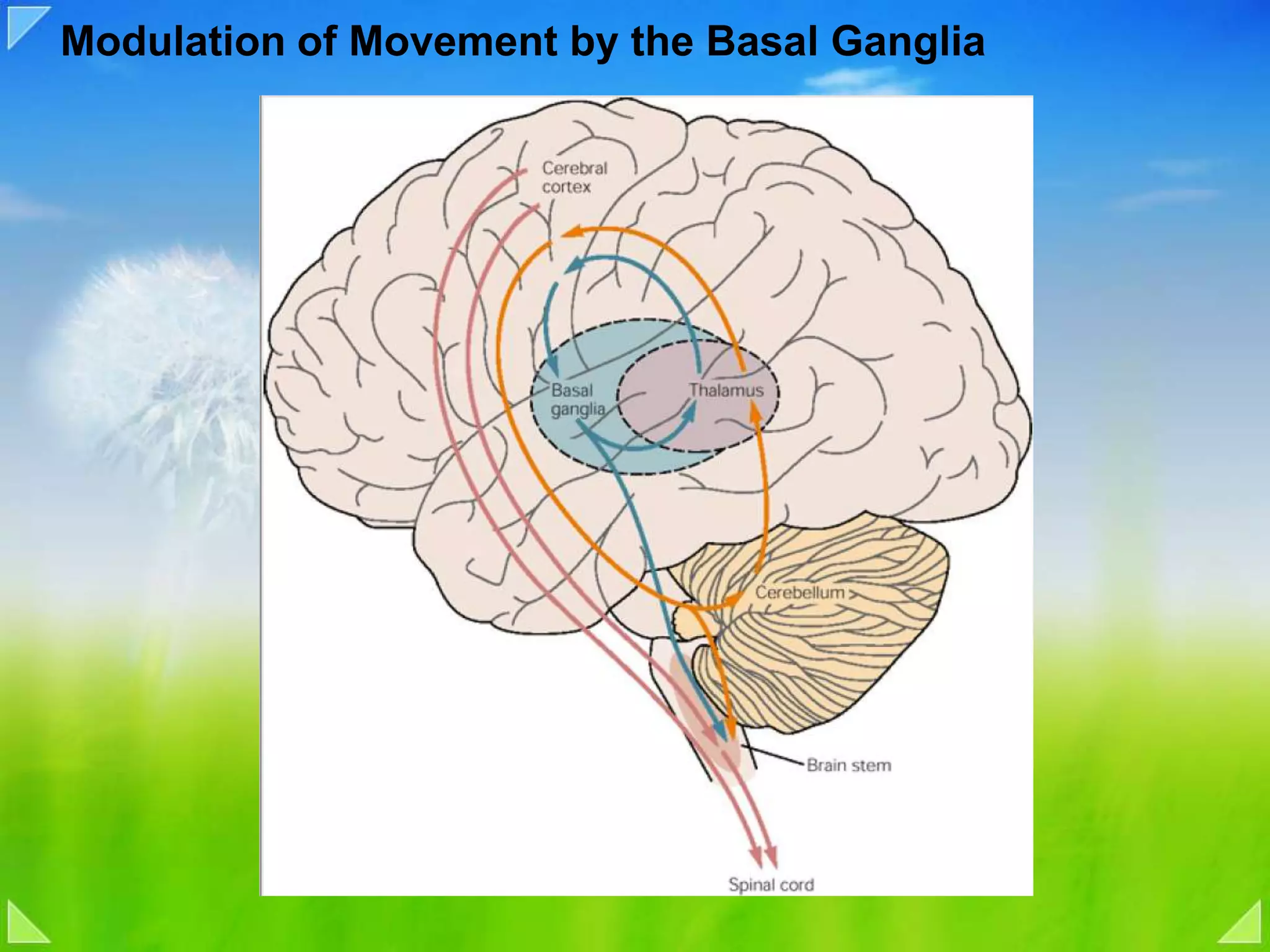
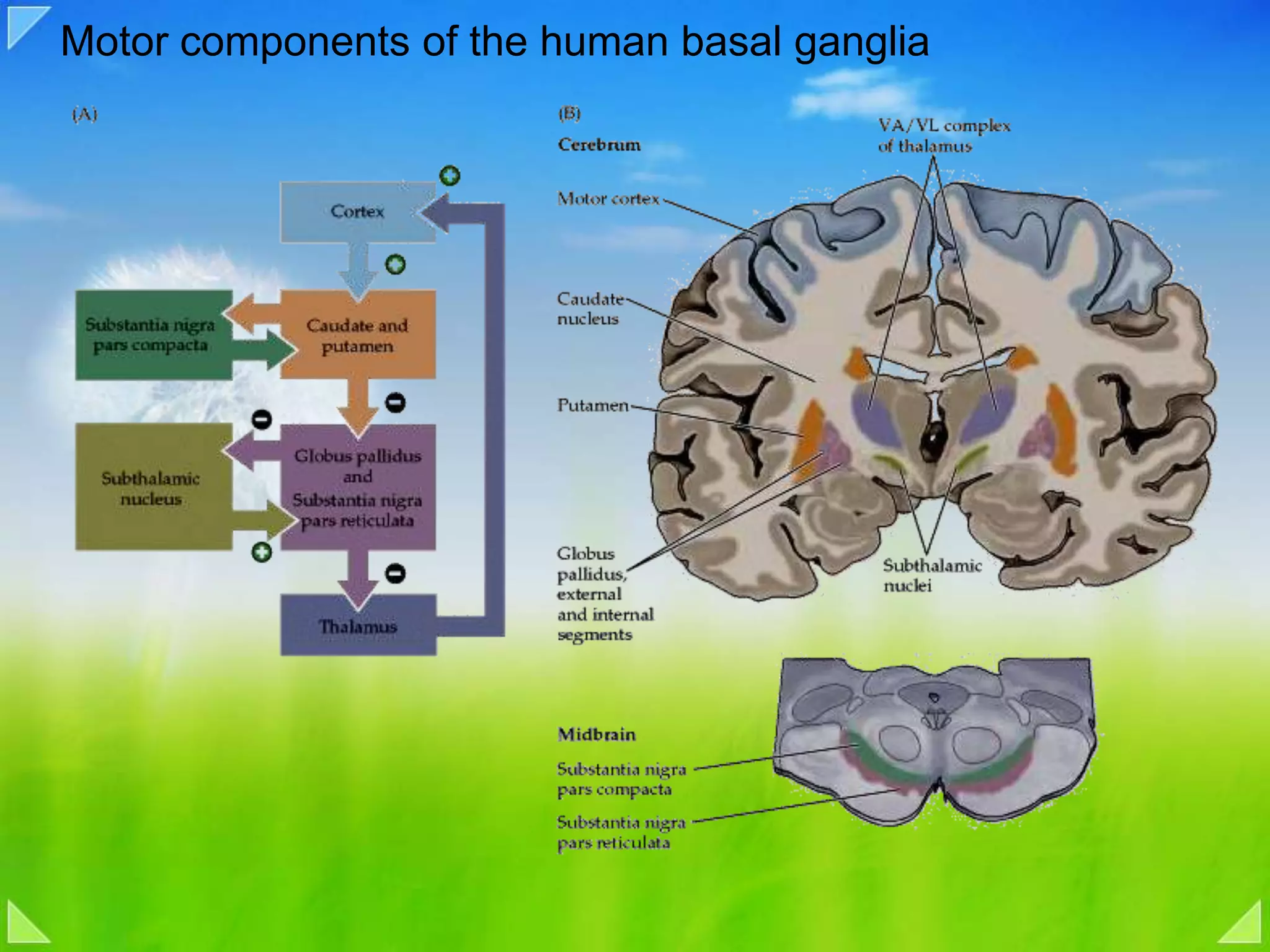
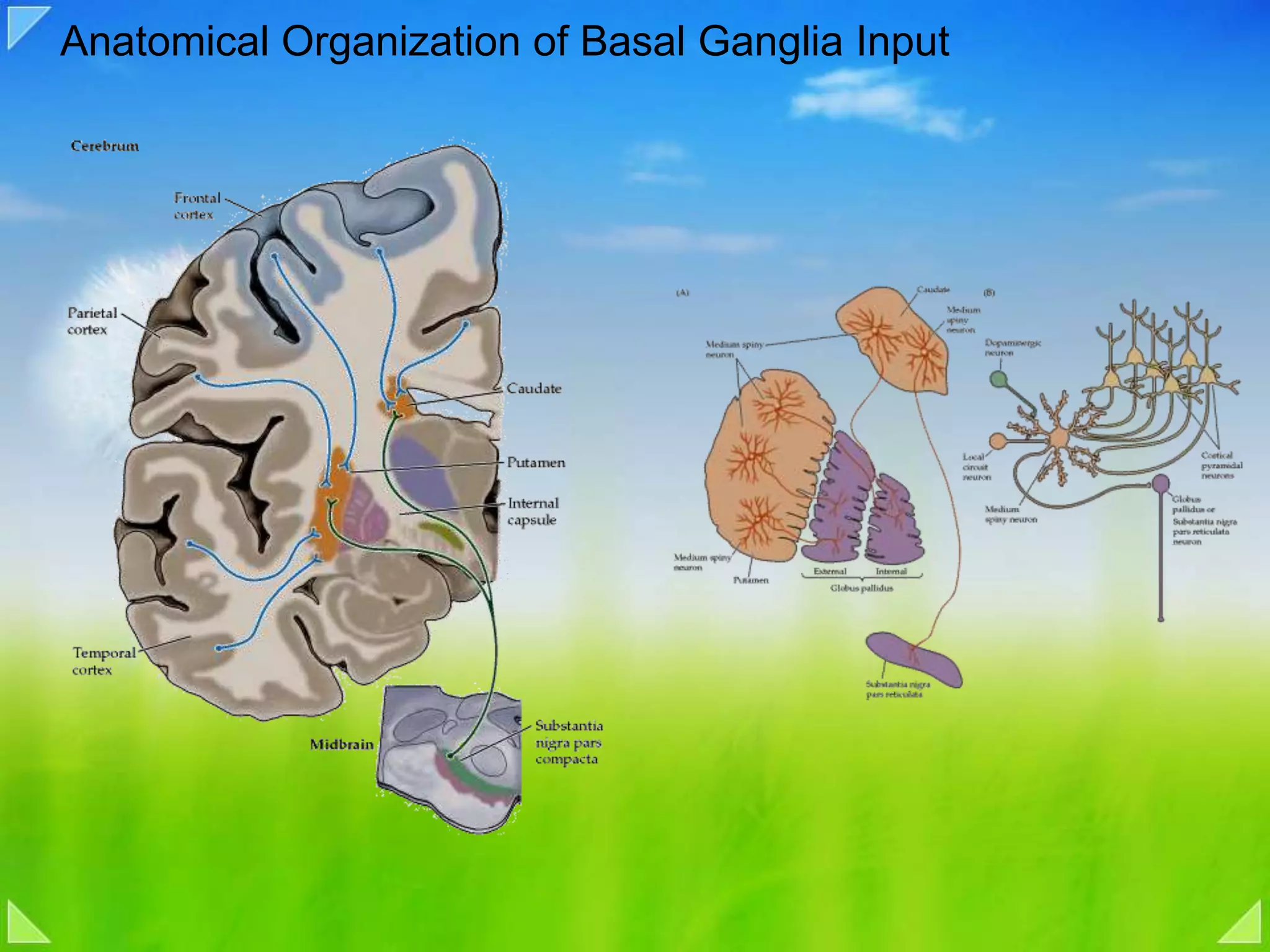
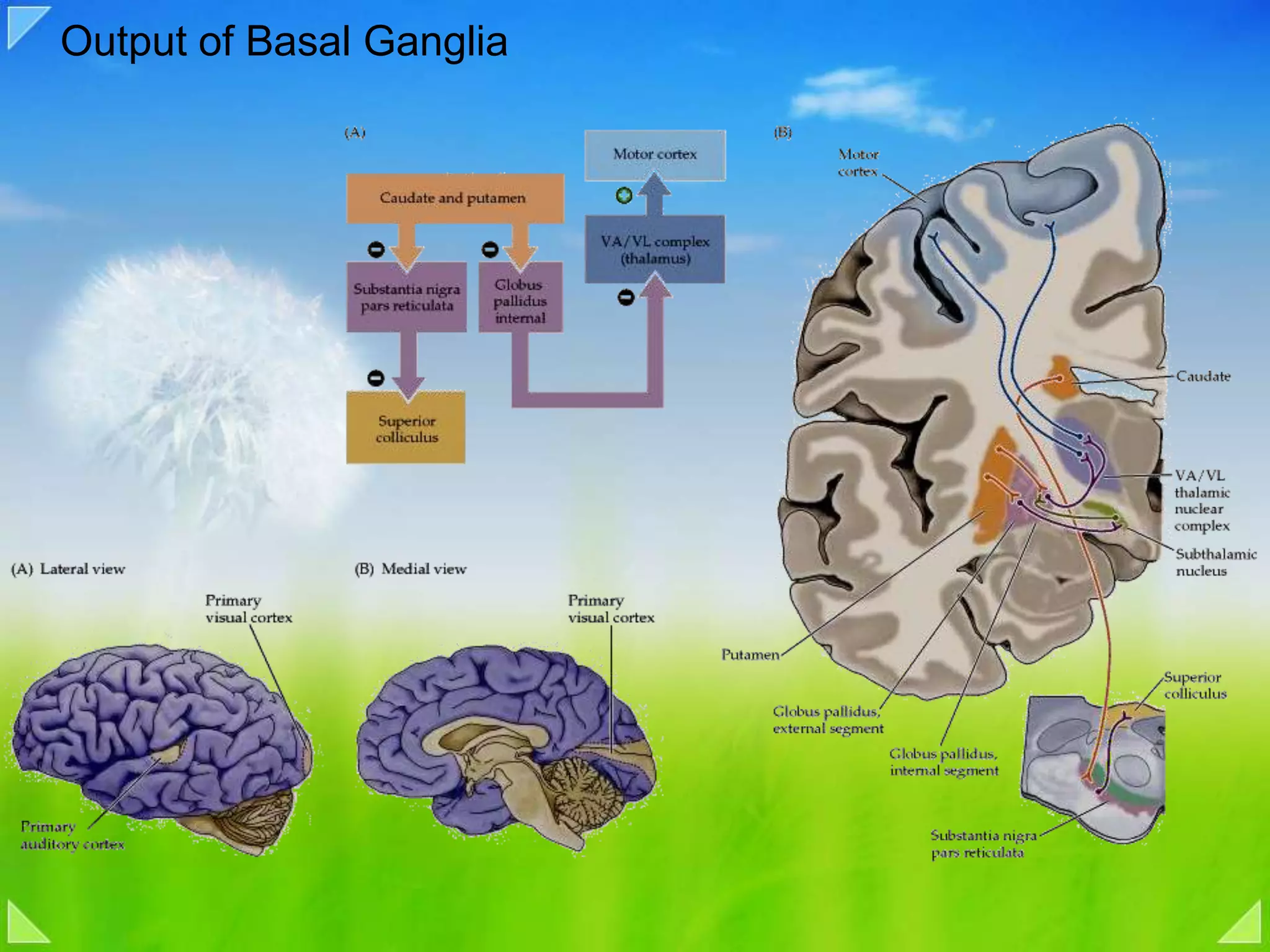
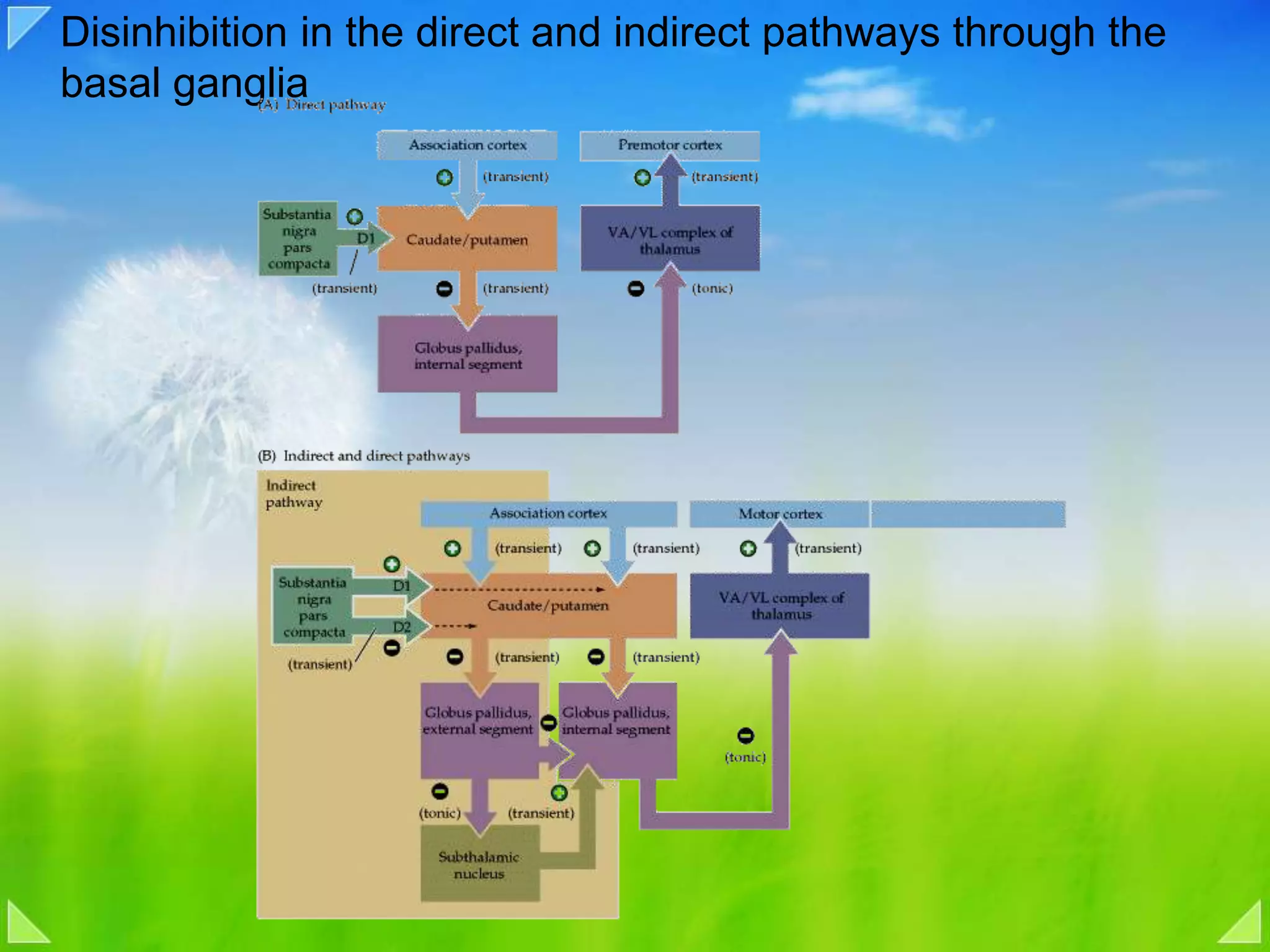
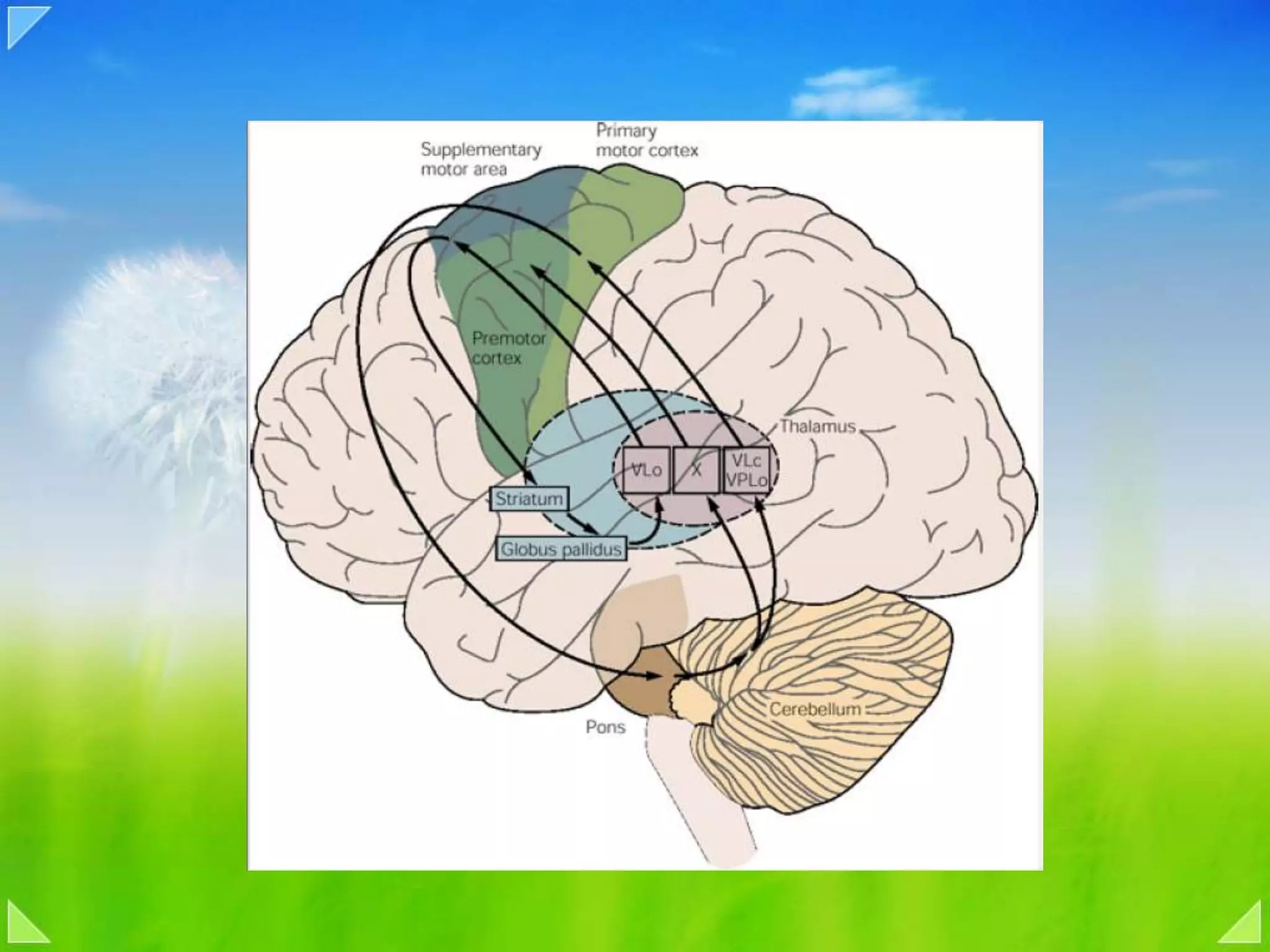
Modulation of movement by the basal ganglia including anatomical structures and pathways.
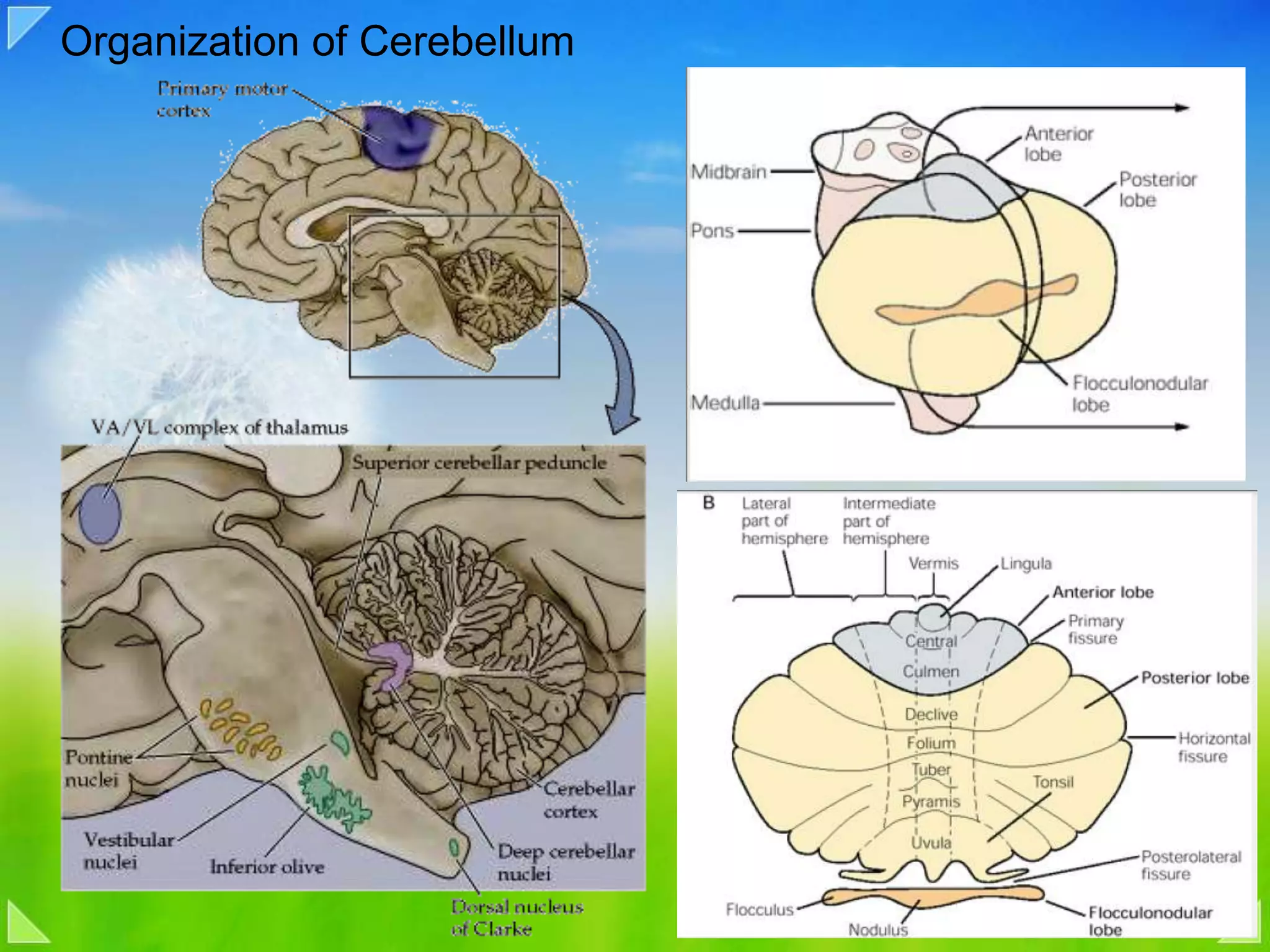
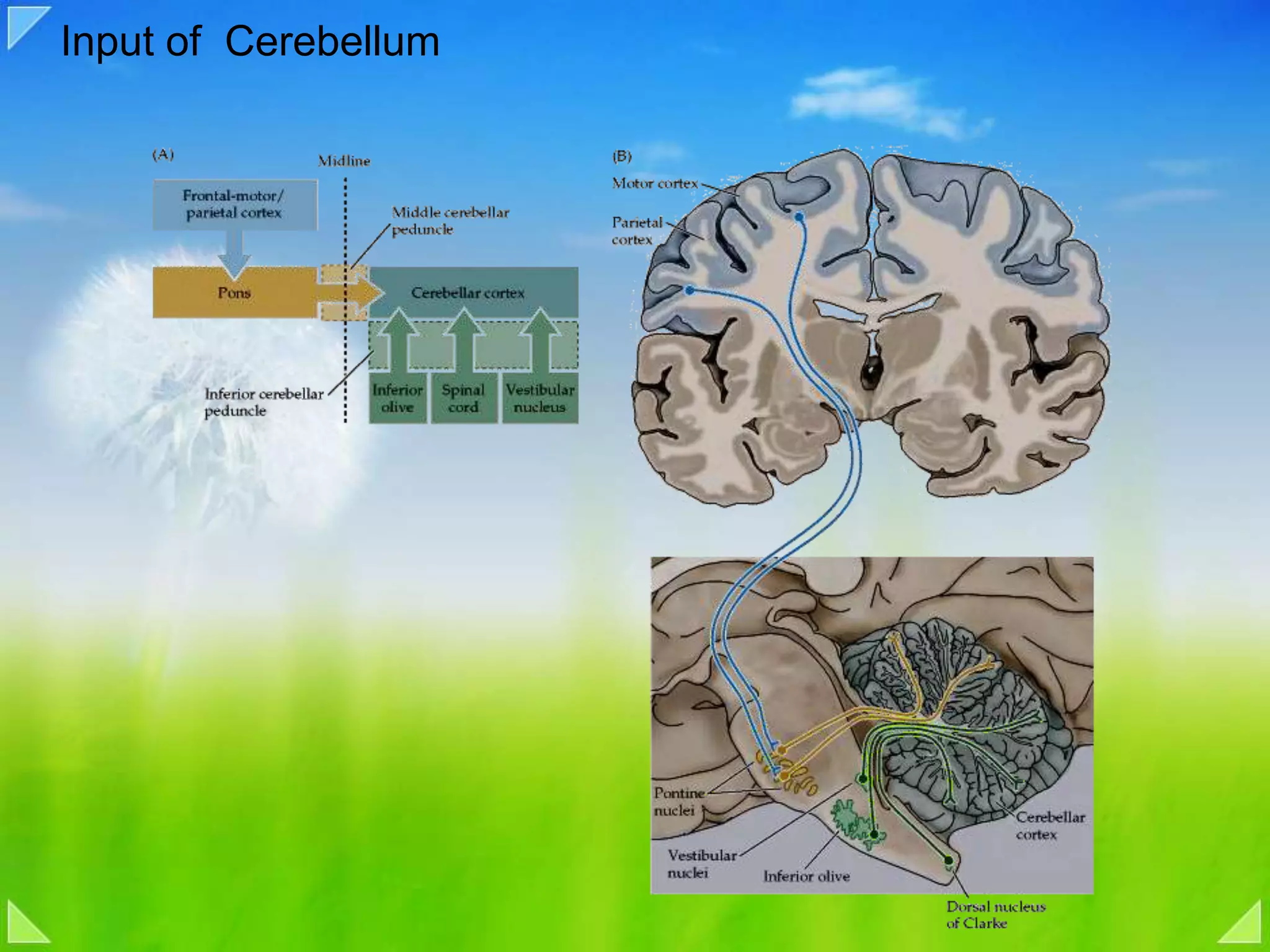
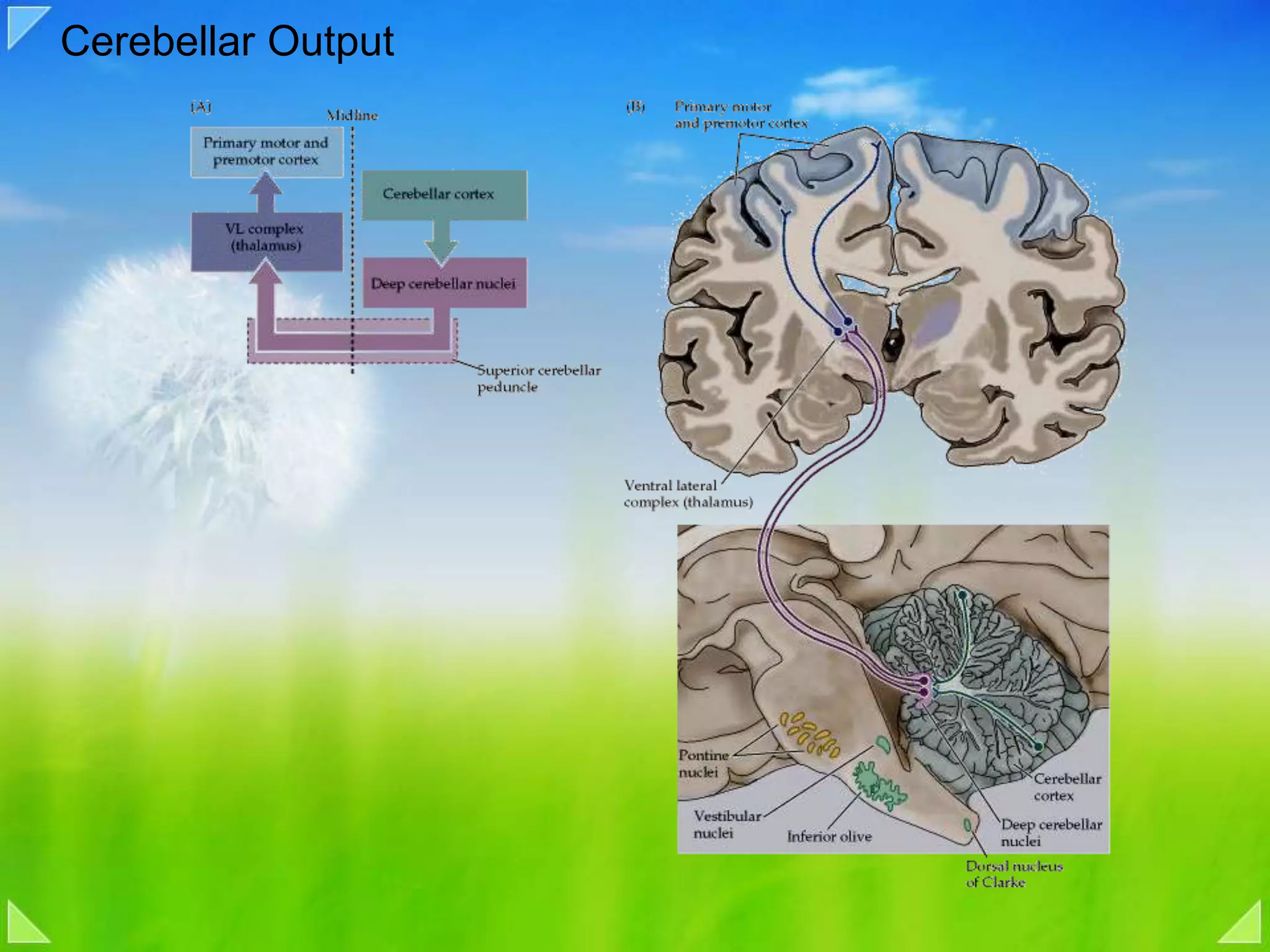
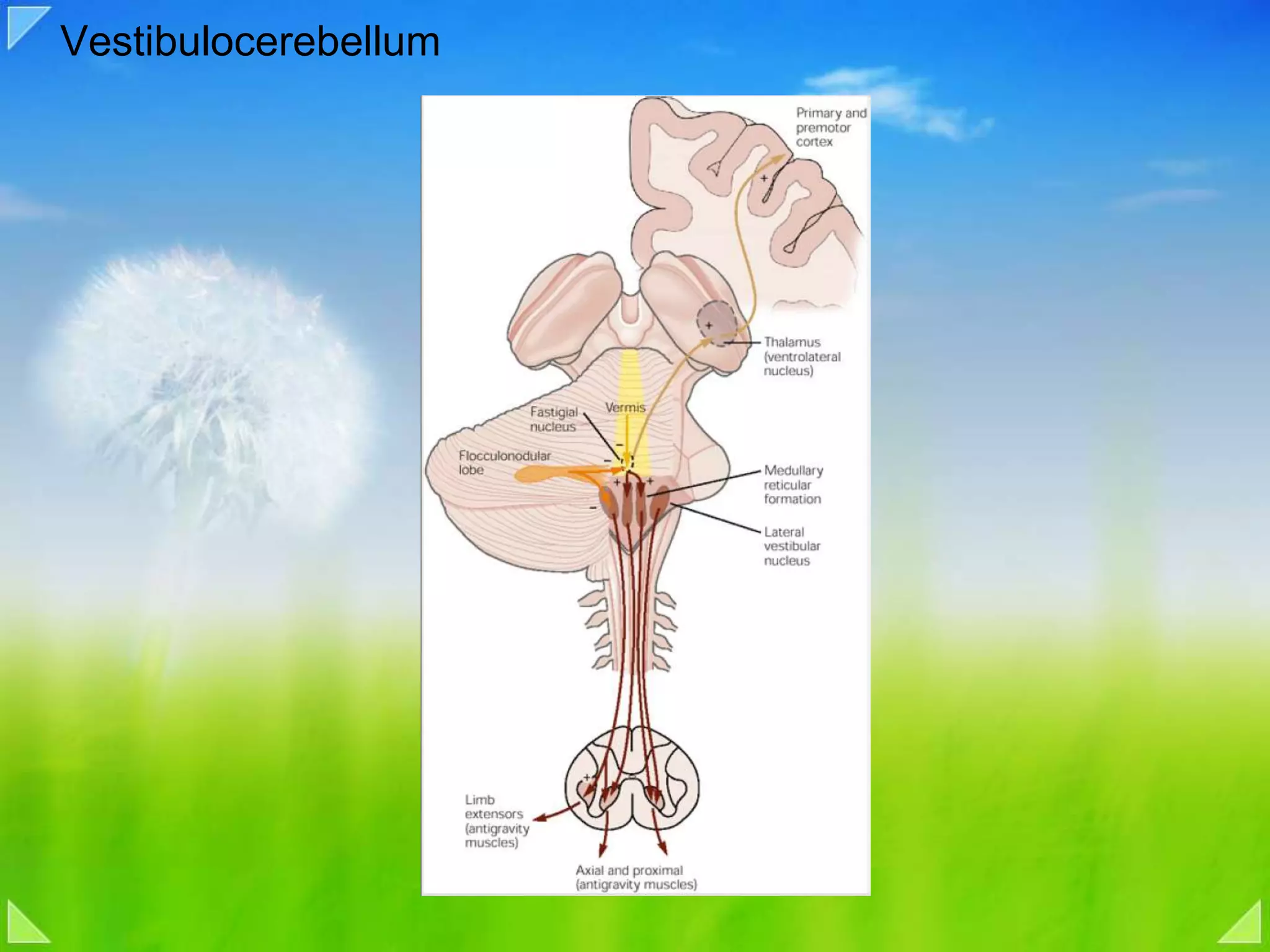
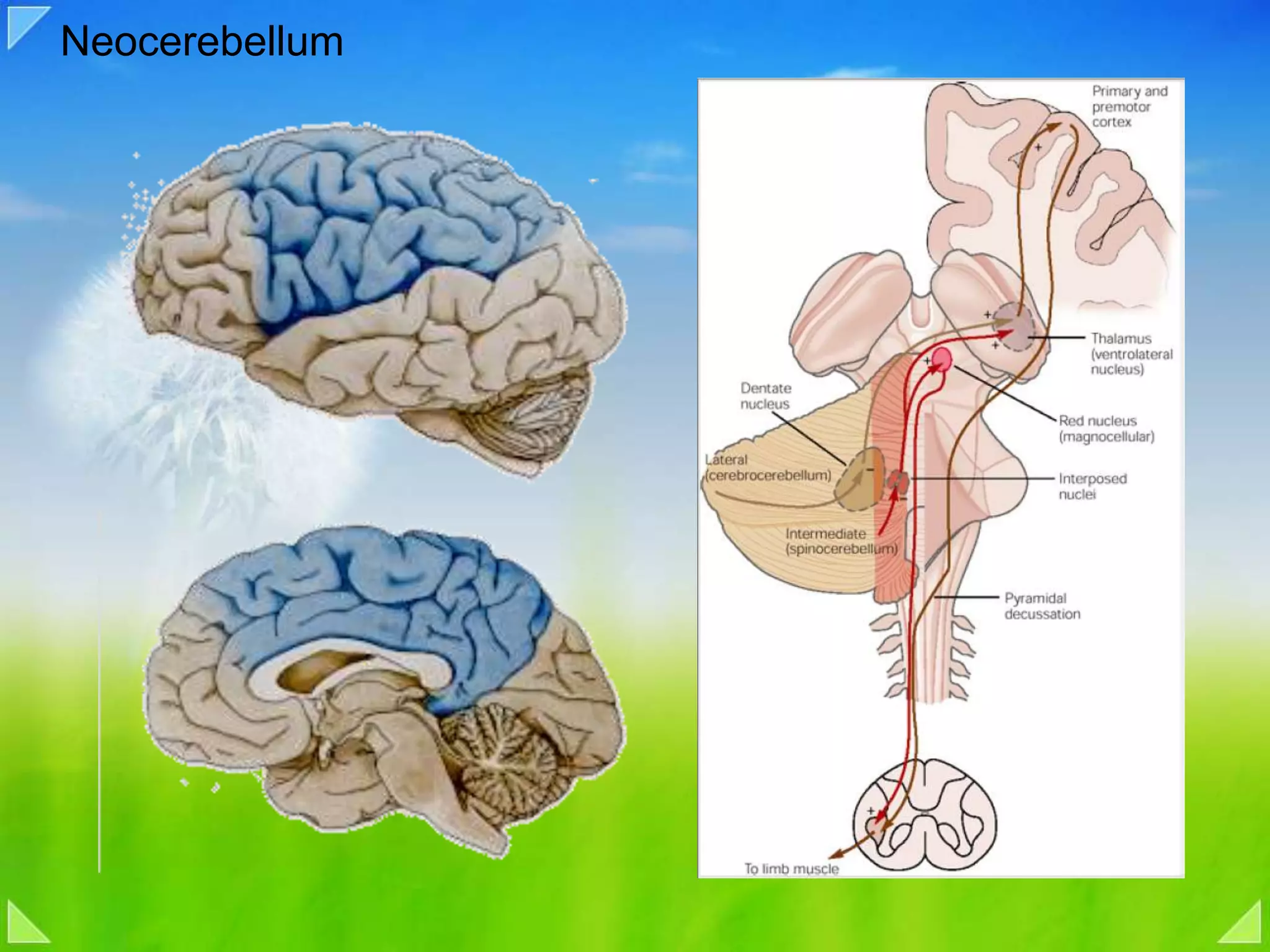
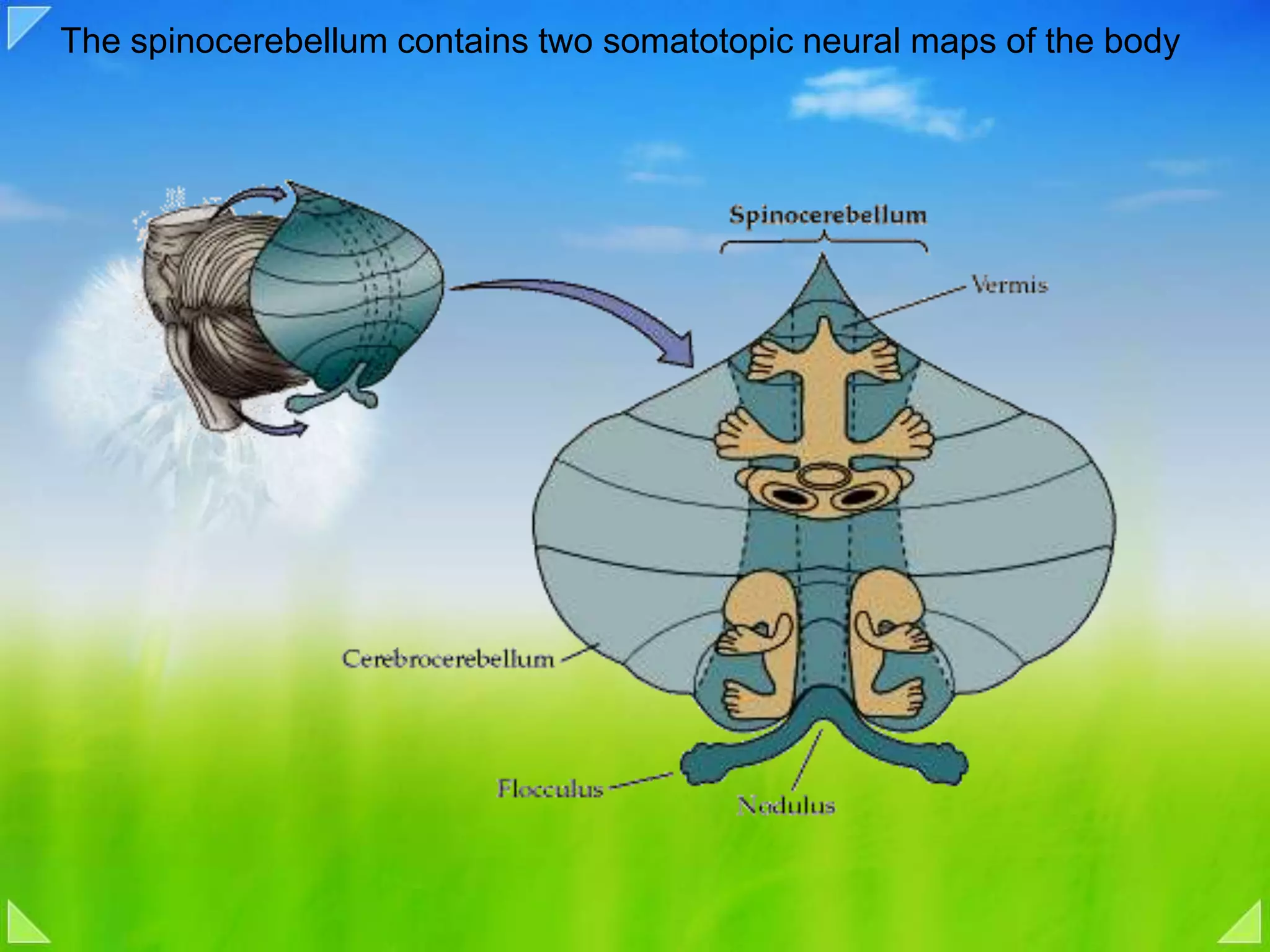
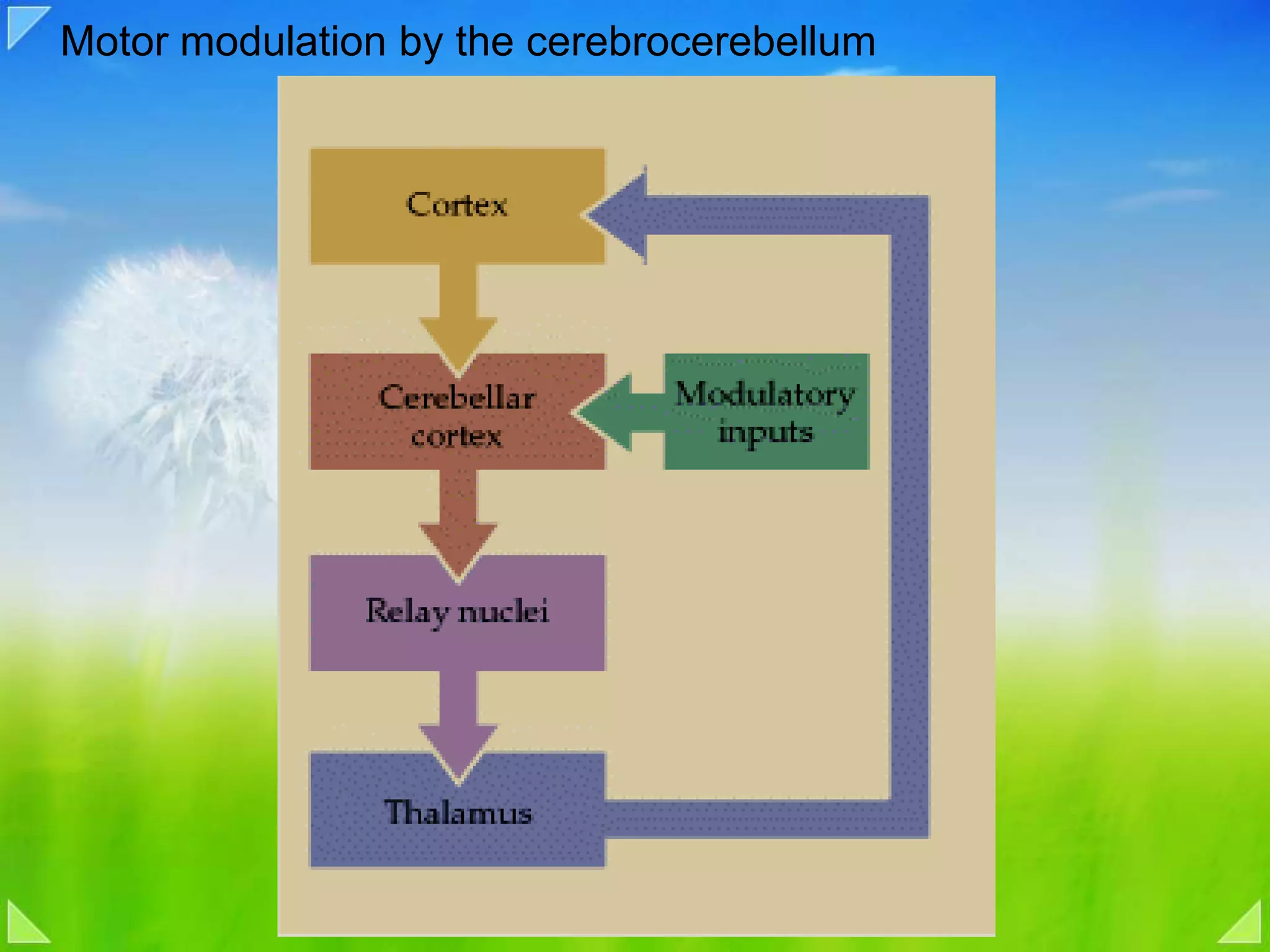
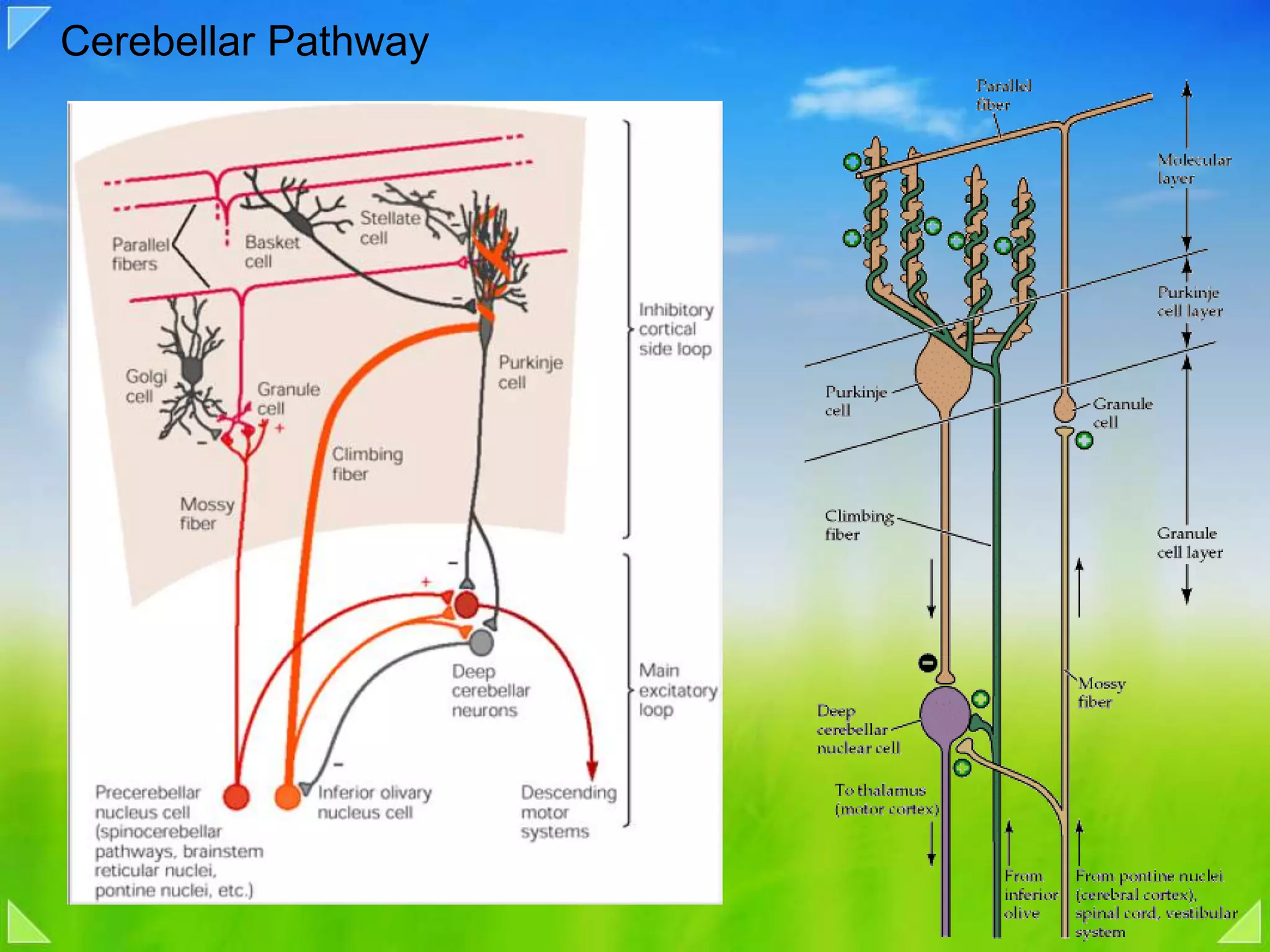
Detailed organizational structures of the cerebellum and its functional outputs.
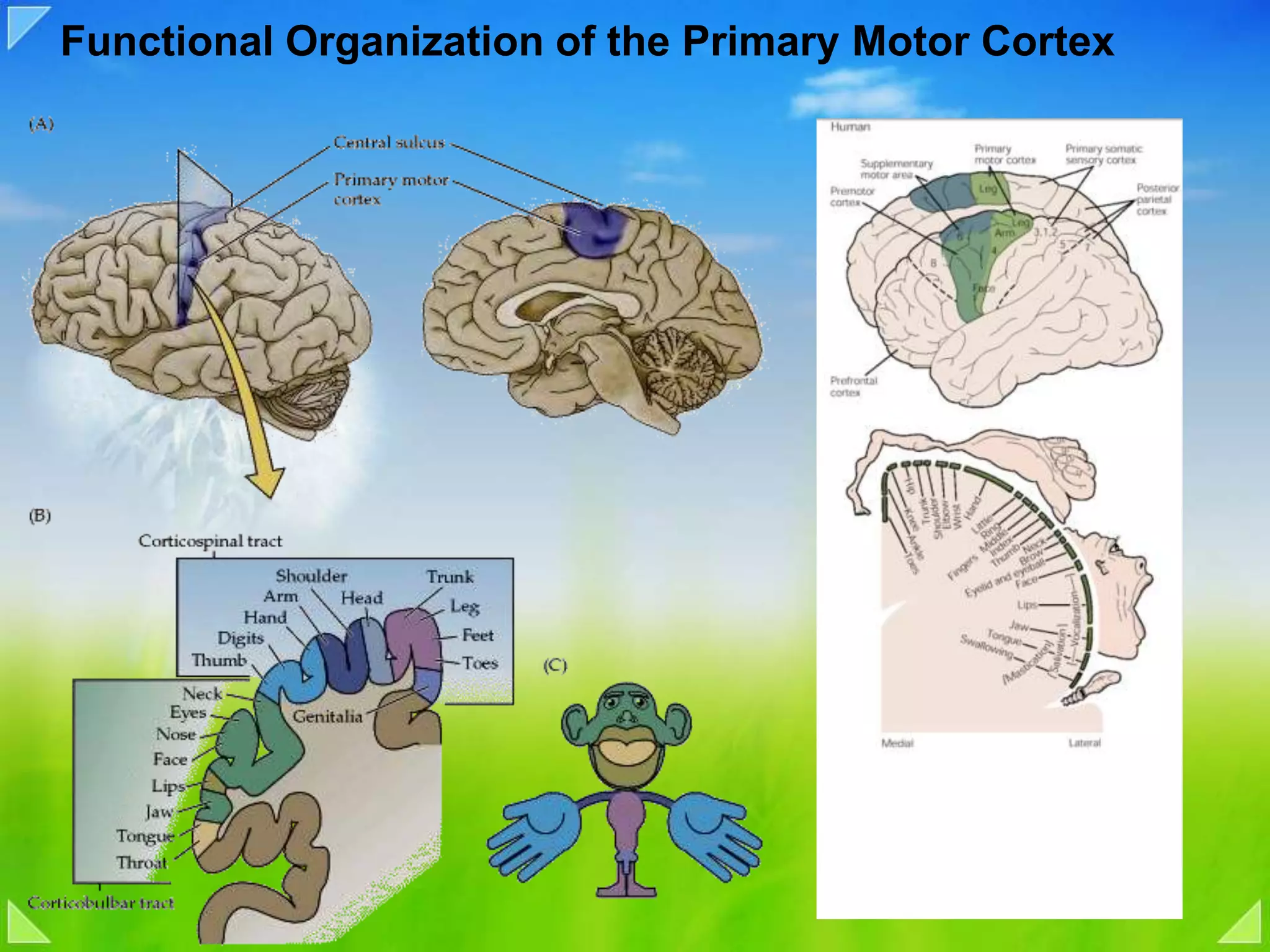
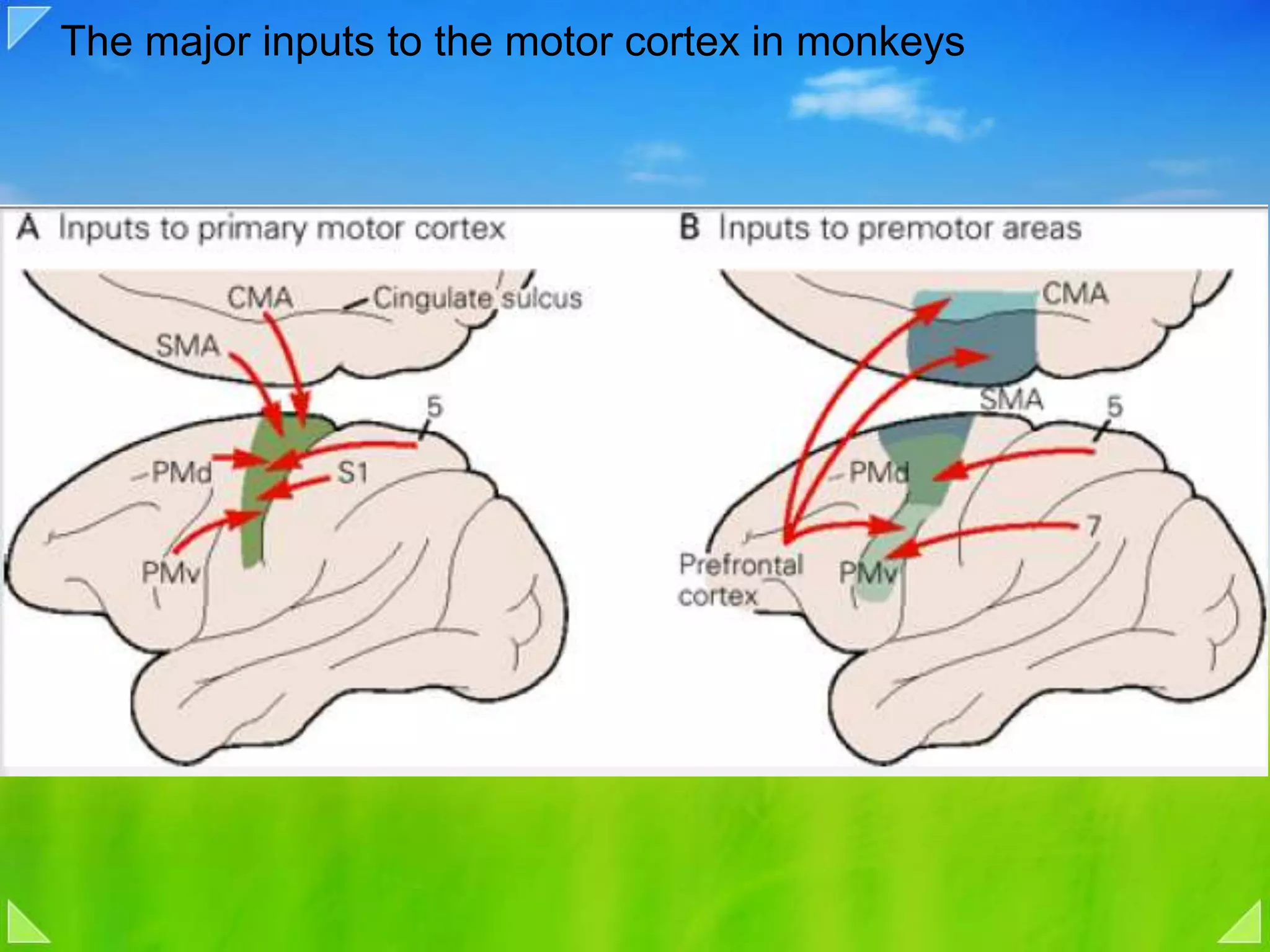
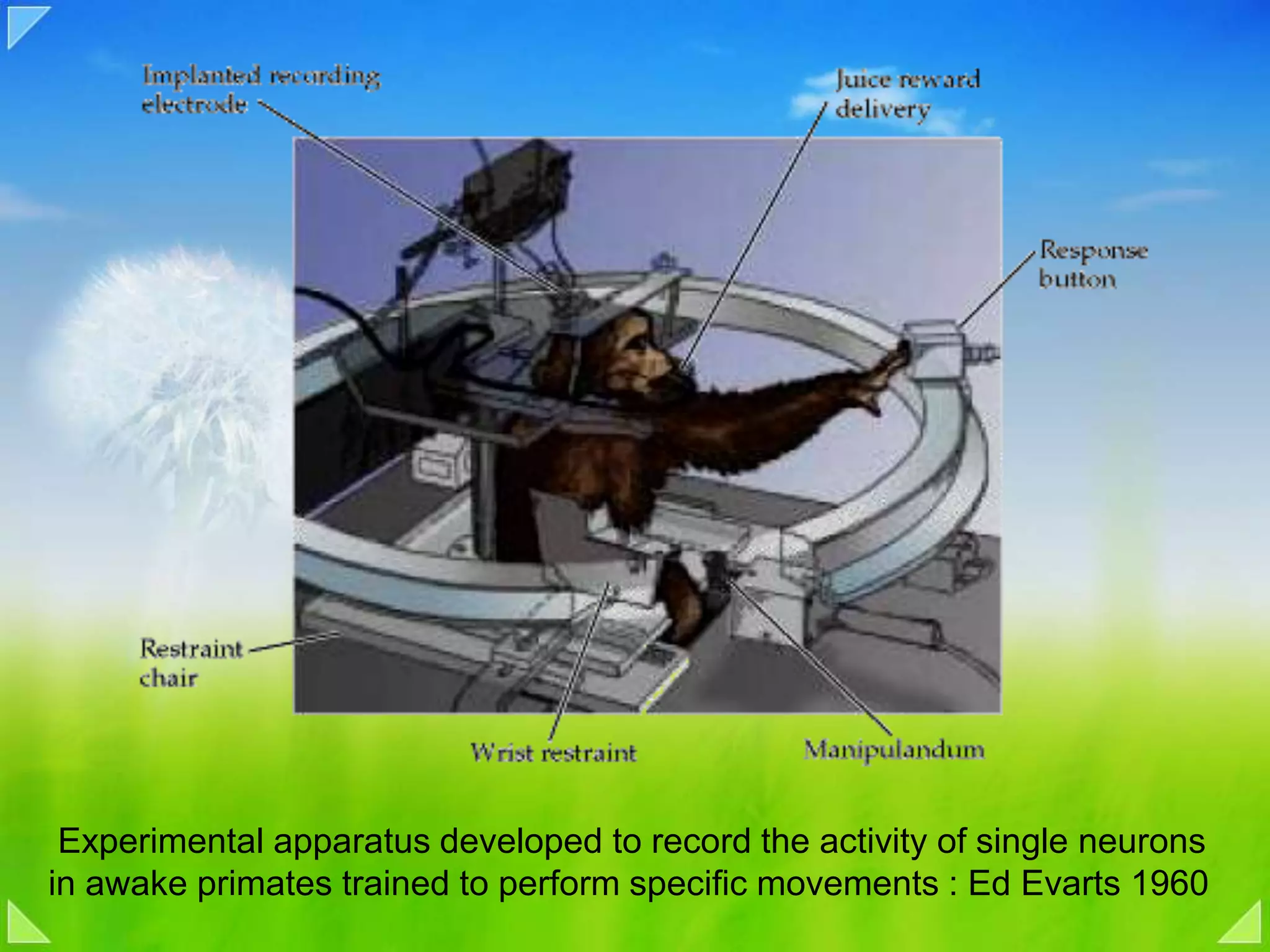
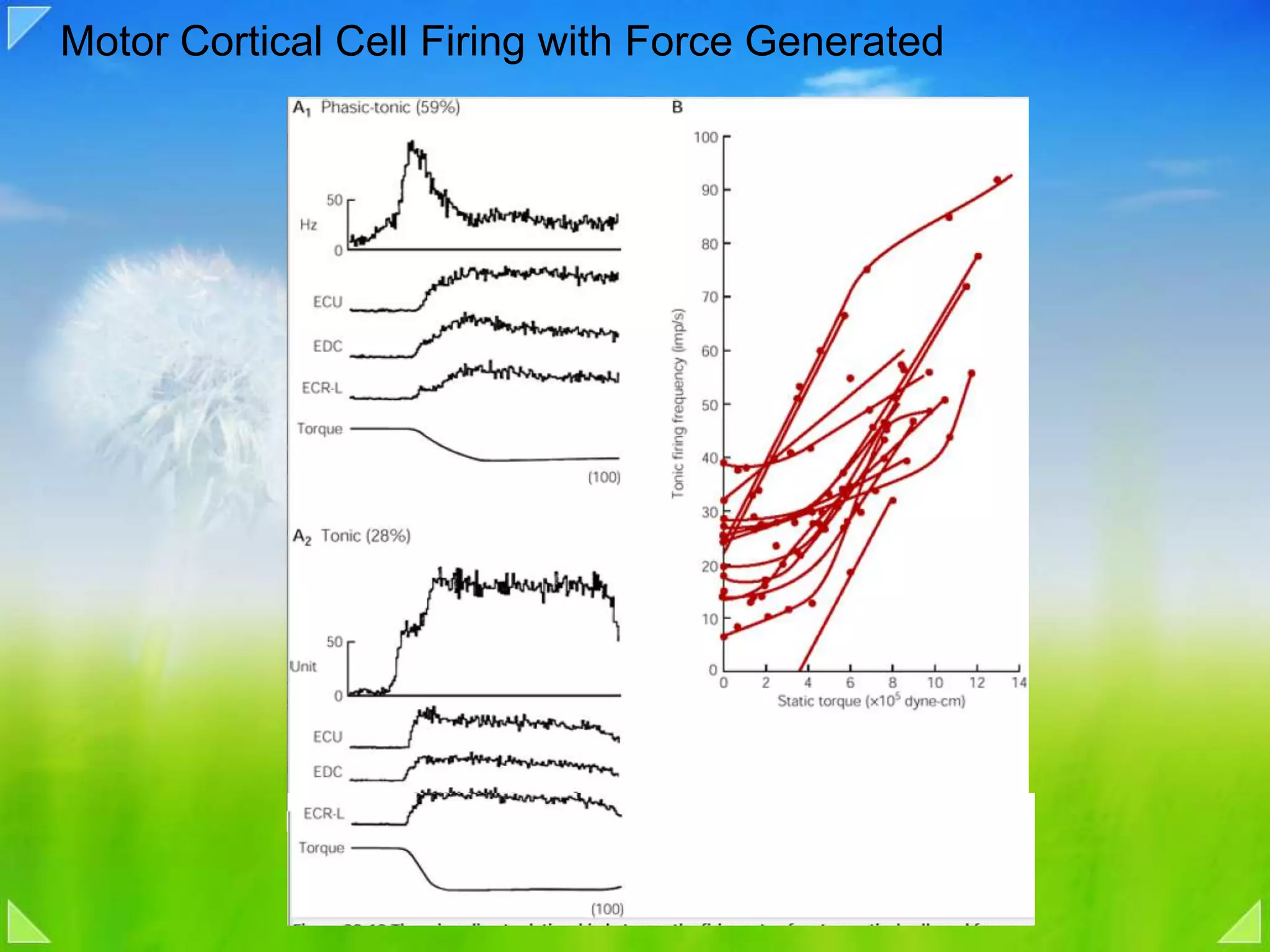
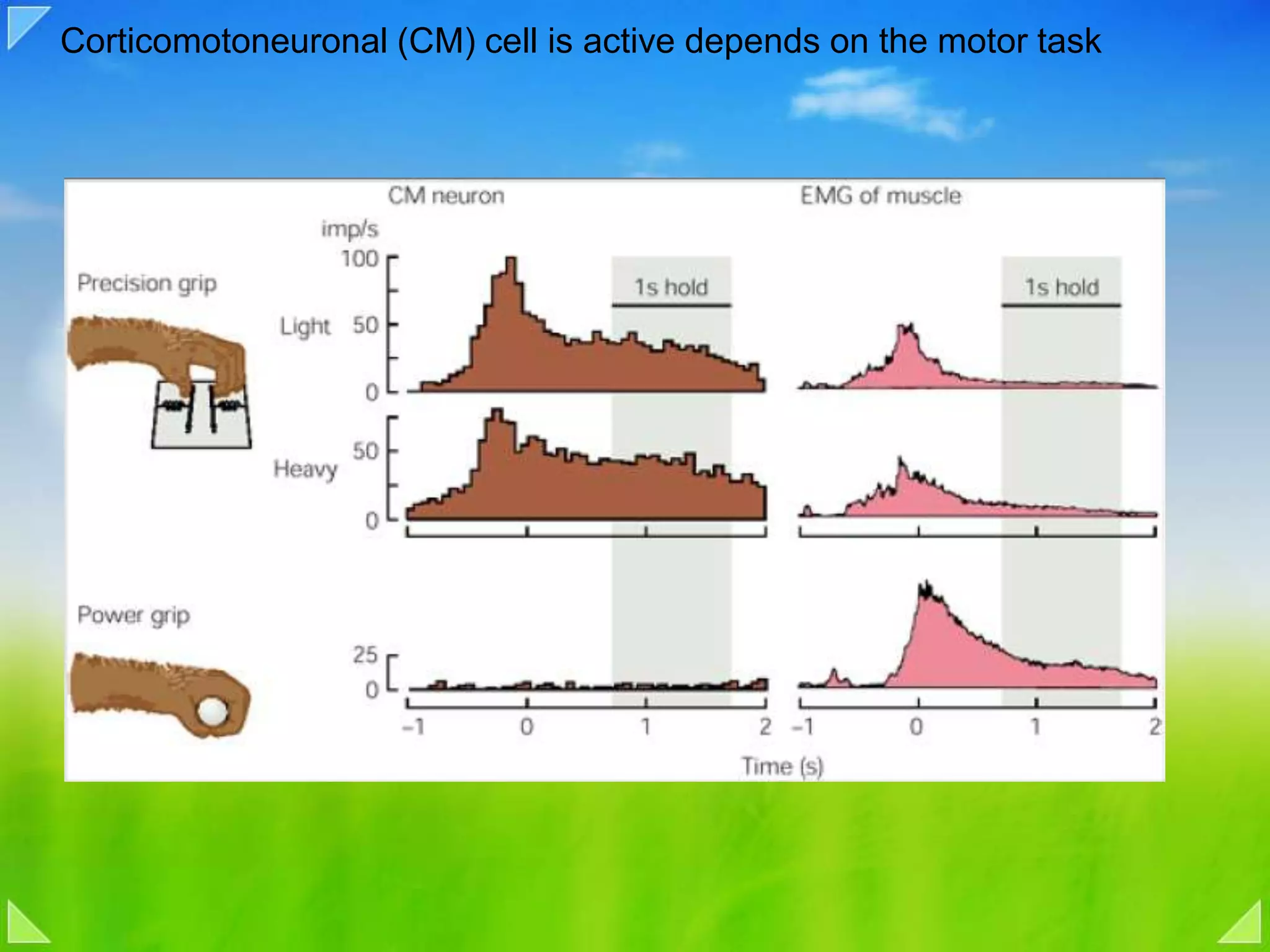
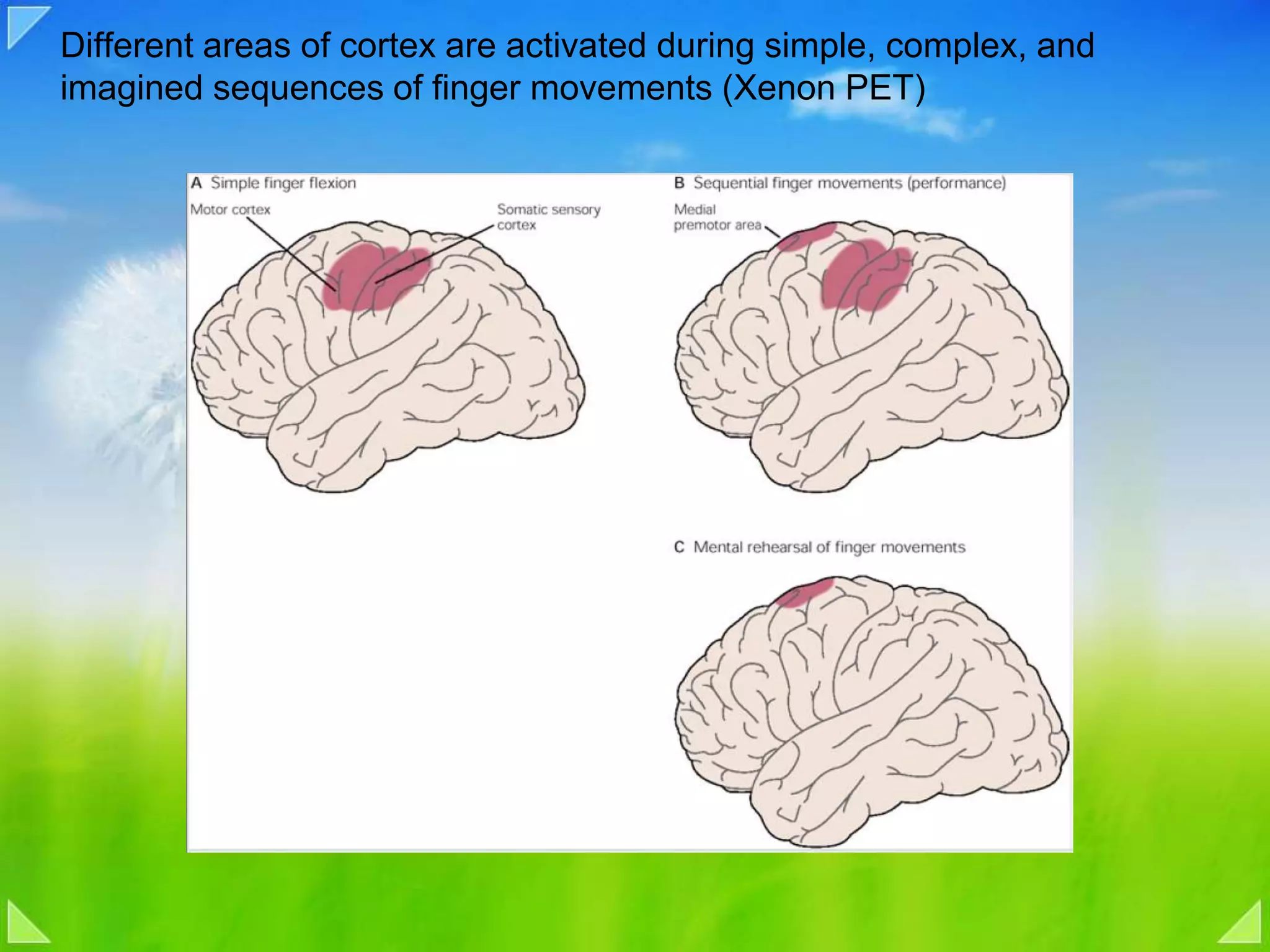
Functional organization of the primary motor cortex and neuronal activity related to movement tasks.
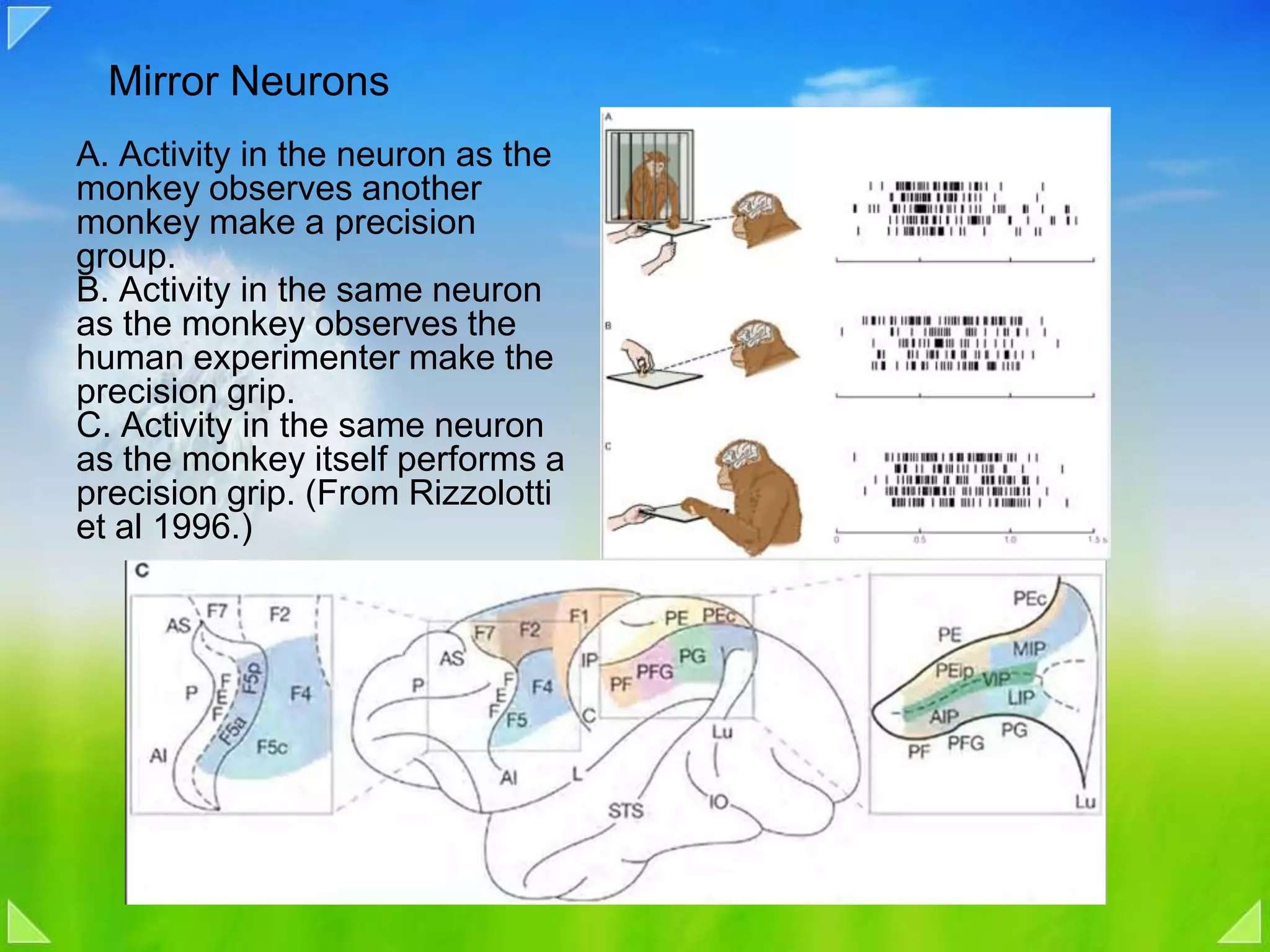
Function of mirror neurons in observing and replicating precision grips.



















































![Share CONTROL OF BODY MOVEMENT[1]PHYSIO PRESENTATION.pptx](https://cdn.slidesharecdn.com/ss_thumbnails/sharecontrolofbodymovement1physiopresentation-250504162954-915fb5e5-thumbnail.jpg?width=640&height=640&fit=bounds)












































