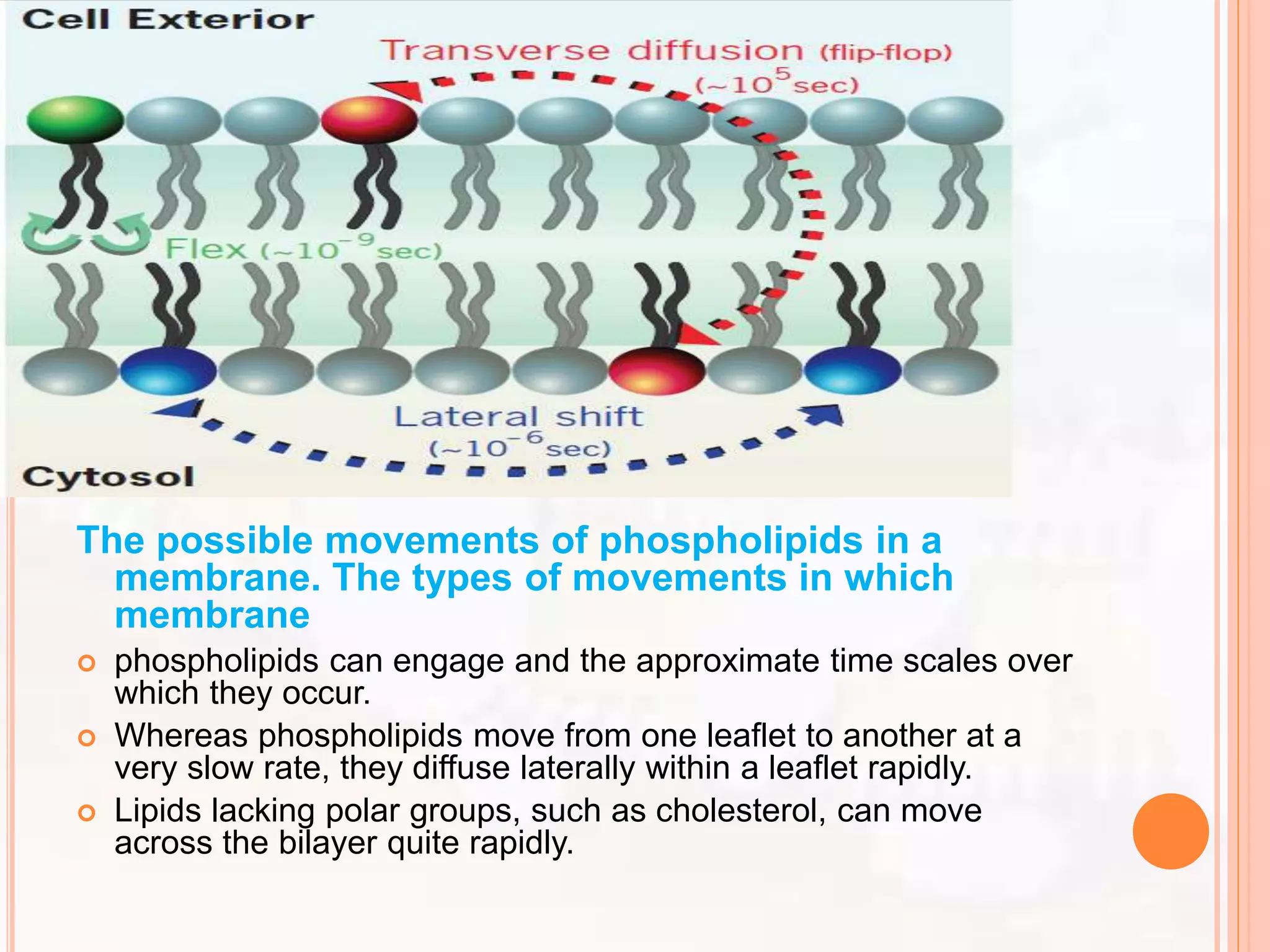
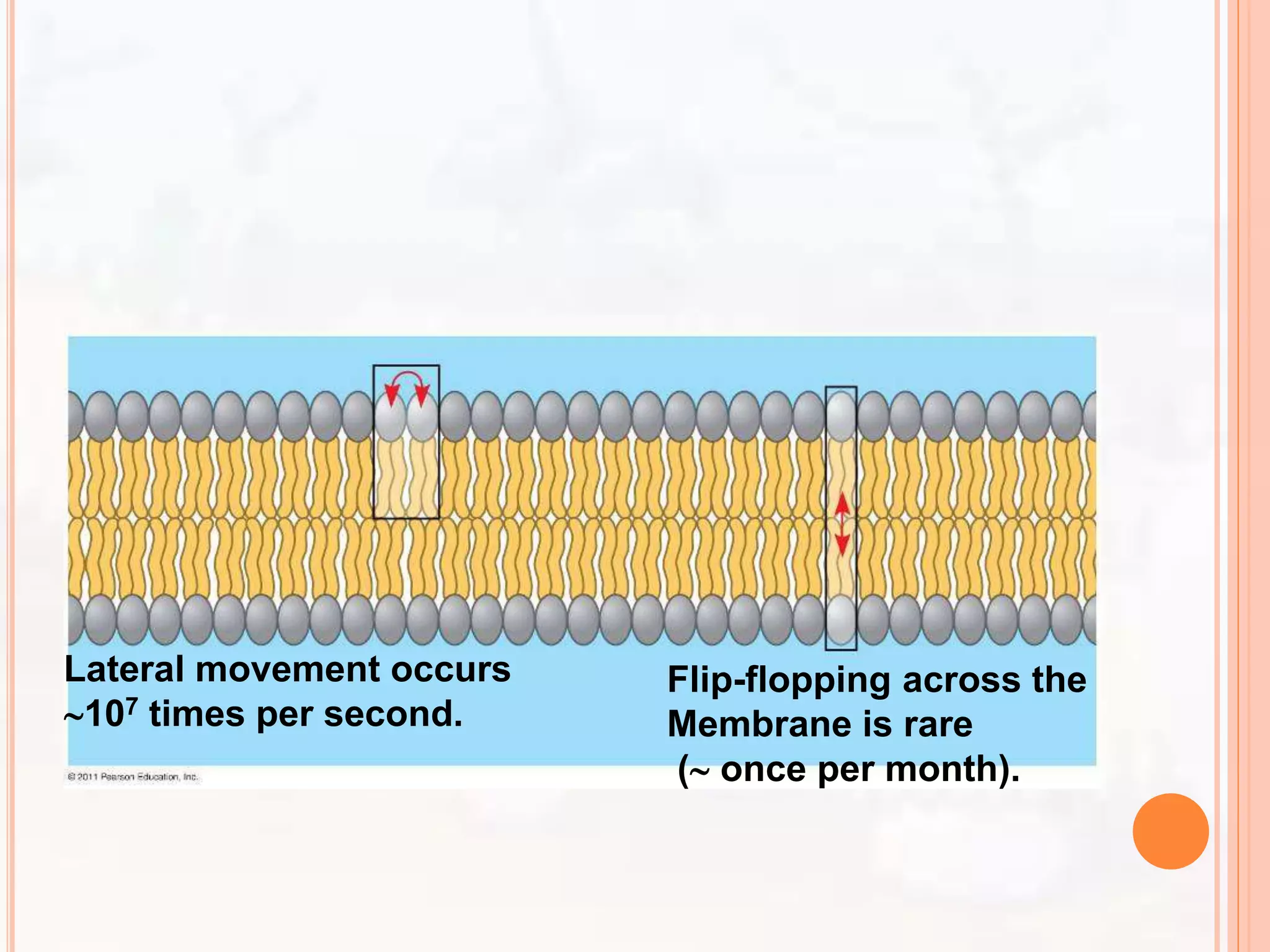
The document summarizes models of the cell membrane structure. It begins by introducing the cell membrane and its functions. It then discusses early models including the lipid bilayer model proposed by Overton, Gorter and Grendel. The protein-lipid-protein hypothesis added proteins to this model. The sandwich model proposed by Danielli and Davson suggested a lipid bilayer sandwiched between protein layers. Robertson's unit membrane model visualized the membrane as a trilaminar structure. Finally, Singer and Nicolson proposed the fluid mosaic model which describes the membrane as a mosaic of lipids and movable proteins embedded within the fluid lipid bilayer.












![……
This basic model has been modified from time to time. Danielli
(1938) suggested the presence of two types of proteins;
tangentially arranged in contact with the lipid and globular
proteins on the outer surface. Again Davson and Danielli
(1943) and Danielli (1954) considered proteins to be in the
form of a folded P-chain.
Perhaps, these units form micelles of membranes indicated in
recent electron micrographs. Membrane models are usually
postulated to contain protein lined polar pores of about 7 Å
diameter which probably permit the passage of small ions and
water molecules across the membrane.
In still other variations the proteins are thought to be in coiled
or globular form on both sides of lipid layers [Fig. 2.3 (A), (B),
(C)] or they are thought to be asymmetrical, with a folded P-
chain on one side and globular proteins on the other [Fig. 2.3
(D)]. Models with globular proteins on both the sides or with
folded P-proteins on both the surfaces and helical proteins
extending into the pores are also suggested.](https://image.slidesharecdn.com/2cellmembrane-221114054551-8d0ce21e/75/cell-membrane-13-2048.jpg)