Download to read offline












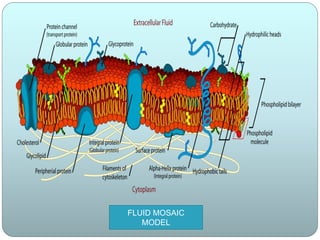

The document discusses cell membranes and summarizes several theories of their structure and function. It begins by describing the basic properties and components of cell membranes. It then summarizes key theories and models of membrane structure proposed by Overton, Langmuir, Gorter and Grendel, Danielli and Davson, Robertson, and Singer and Nicolson. Their fluid mosaic model from 1972 proposed that membranes are a fluid bilayer of lipids with integral and peripheral proteins dispersed within.











































































