Downloaded 79 times
































![Neopilina galatheae , top (dorsal) view. Neopilina is peculiar because of the replication of various of its organs and organ systems, reminiscent of metameric animals. The class Monoplacophora is well known as fossils, and until 1952 all of its members were believed to have been extinct since the Devonian period, about 350 million years ago. [This shell, relatively thin, was damaged while being dredged.]](https://image.slidesharecdn.com/chapter-12-1208567734985791-8/85/Chapter-12-33-320.jpg)
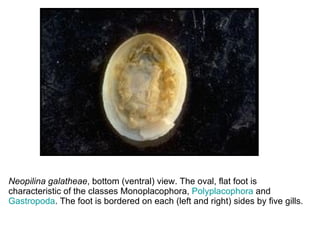
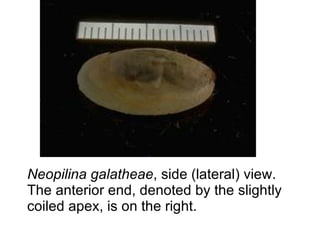



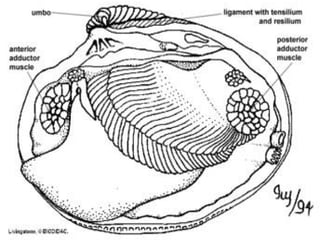
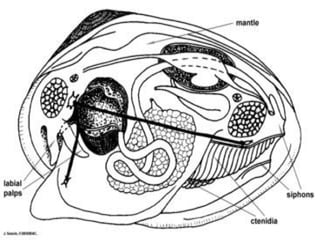
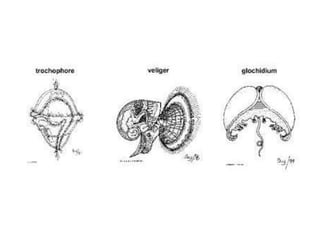
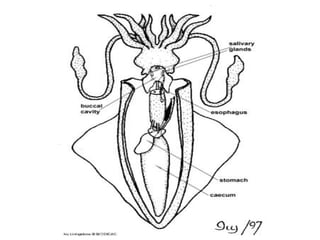
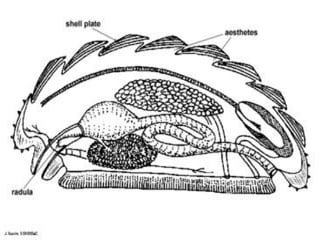
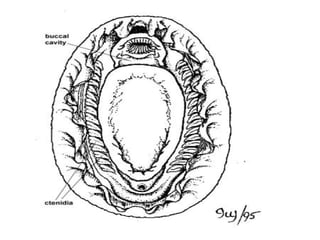


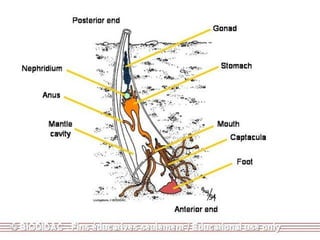

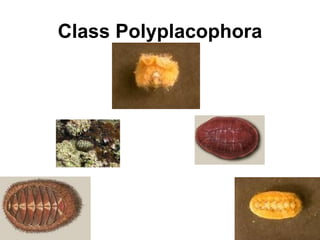
The document discusses the characteristics of the mollusca phylum. It notes that molluscs have soft bodies and come in a wide variety of forms, including snails, clams, and octopuses. The phylum is very successful, with over 100,000 species, and molluscs have adapted to live in many different environments through evolution. The three main classes of molluscs - gastropods, bivalves, and cephalopods - are also introduced.
























































![Mollusks%20bivalves%3agastro[1]](https://cdn.slidesharecdn.com/ss_thumbnails/mollusks20bivalves3agastro1-120203103120-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)















































