
Downloaded 40 times







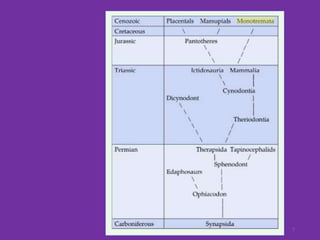

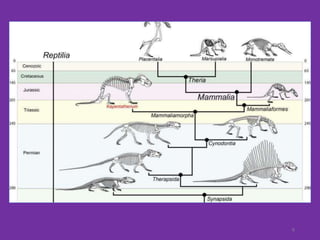
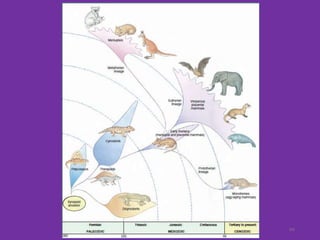
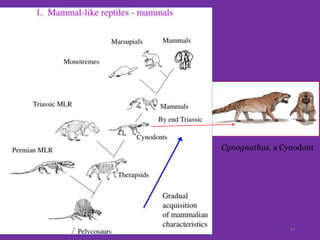







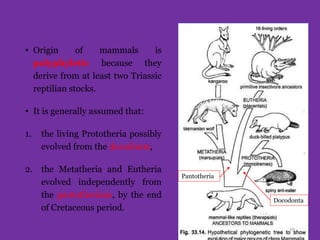





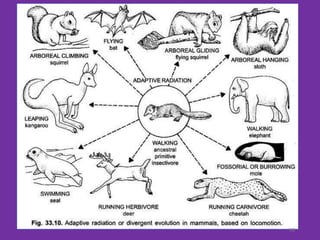











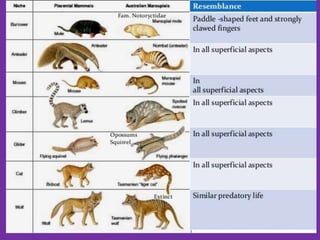

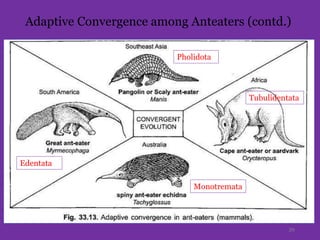
The document discusses the origin and evolution of mammals. It describes two main theories for the ancestry of mammals - through amphibians or reptiles. While amphibian ancestry was proposed, reptilian ancestry is now widely accepted based on fossil evidence. Many characteristics of early mammal-like reptiles called therapsids were mammalian. True mammals first appeared in the Jurassic period but remained small until after the extinction of dinosaurs. When ecological niches opened up in the Cenozoic era, mammals underwent adaptive radiations into various forms through modifications of limbs, teeth and other features for different habitats like trees, ground, burrows, water and air. Convergent evolution also led to similarities between unrelated mammals adapting to the same nic























































































