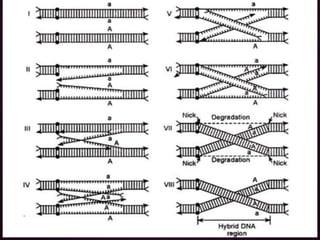
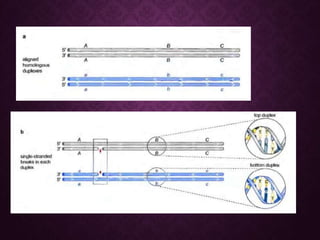
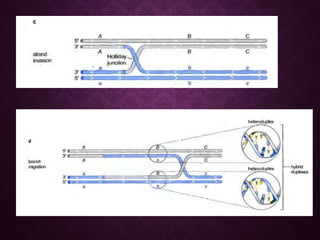
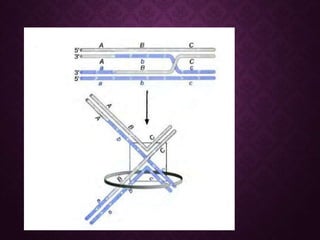
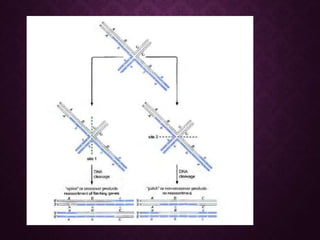
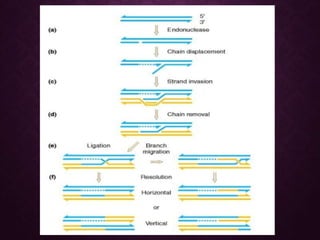
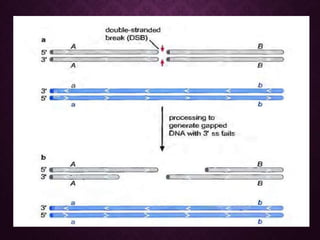
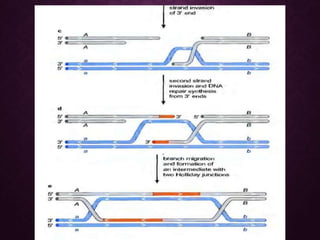
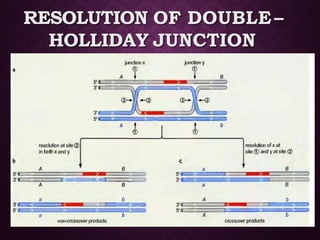
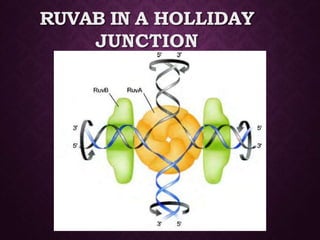
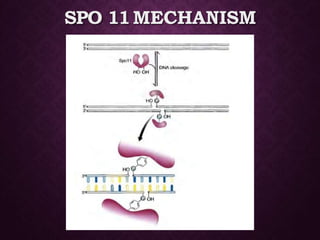
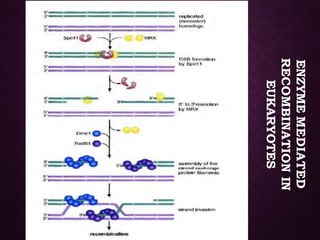
This document discusses molecular genetics topics including sex determination, genetic recombination, and transposons. It outlines four units of study: 1) Sex determination and dosage compensation, 2) Genetic recombination mechanisms and models, 3) Enzymes involved in homologous and site-specific recombination, and 4) Bacterial and eukaryotic transposons. The second section provides detailed descriptions of homologous recombination models including Holliday, Whitehouse, Meselson-Radding, and double-strand break repair pathways. Key enzymes in E. coli recombination such as RecBCD, RecA, RuvAB, and RuvC are also summarized.




































































































![Polymer [ बहुलक ] Chemistry Notes PDF - Irfanullah Mehar - JJ Sir Chemistry.pdf](https://cdn.slidesharecdn.com/ss_thumbnails/polymerchemistrynotespdf-irfanullahmehar-jjsirchemistry-260210172118-3f9b37f7-thumbnail.jpg?width=640&height=640&fit=bounds)














