The document discusses leaf growth and differentiation, outlining the stages of leaf ontogeny from initiation at the shoot apical meristem (SAM) to the completion of histogenesis. It details the role of genetic factors, including the plastochron gene in rice and various homeobox genes, in regulating leaf formation and development. The mechanisms of auxin transport and gene interactions that influence leaf primordium initiation are highlighted, emphasizing the complex regulatory networks involved in leaf morphology.












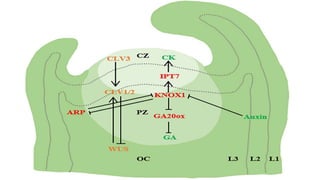
![• SAM is divided into three functional regions [central region (CZ),
peripheral region (PZ) and costal region (OC)] and Layer1 (L1), Layer2
(L2) and Layer3(L3). WUS activates CLV3, and CLV3 further binds to
CLV1/2, thereby inhibiting WUS expression. Auxin accumulation in the
flanks of SAM through PIN1/AUX1 mediated polar transport triggers
primordium development. In addition, KNOX1 maintains the role of stem
cells, positively regulates CK, negatively regulates GA signaling through
IPT7 and GA20ox, and ARF regulates the emergence of young primordia.](https://image.slidesharecdn.com/leafinitiationgrowthanddifferentiation-240603111107-1ab5884b/85/Leaf-Initiation-Growth-and-Differentiation-pdf-16-320.jpg)

![• The second mechanism is the mutual inhibition of the tip meristem maintenance
gene KNOX1 and ARP [ASYMMETRIC LEAVES1(AS1)/ROUGH
SHEATH2(RS2)/PHANTASTICA (ARP)].
• The downregulation of KNOTTED1 ( KN1 ) class of homeobox-containing plant
genes that were originally identified in maize in fact provided an early molecular
marker of leaf initiation in SAM (Brutnel and Langdale 1998 ; Sinha 1999 ).](https://image.slidesharecdn.com/leafinitiationgrowthanddifferentiation-240603111107-1ab5884b/85/Leaf-Initiation-Growth-and-Differentiation-pdf-18-320.jpg)


















































































