Download as PDF, PPTX











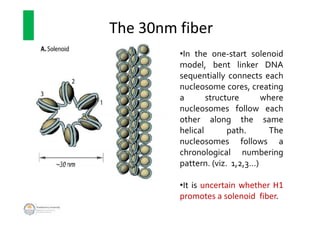





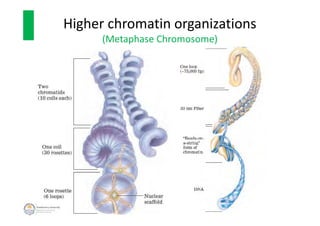









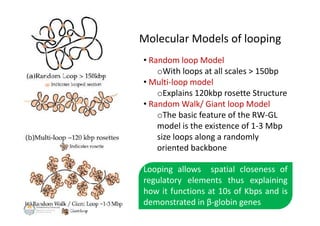








The document discusses genome organization in eukaryotes. It describes how DNA is highly condensed and packaged within the nucleus through different levels of organization, from nucleosomes to 30nm fibers and higher-order structures. DNA is wrapped around histone proteins to form nucleosomes, which further condense into 30nm fibers. These fibers compact to form loops, domains, and chromosome territories within the nucleus. The precise structures at higher levels of organization are still being elucidated. Precise packaging is necessary to condense the large eukaryotic genome while allowing access for processes like transcription and replication.























































































