



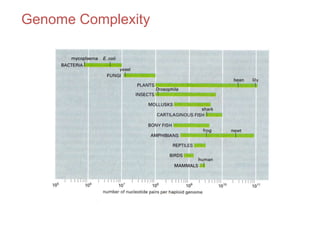

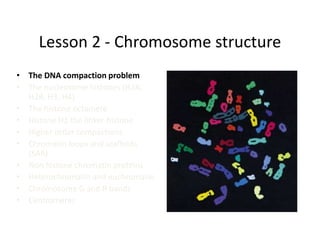







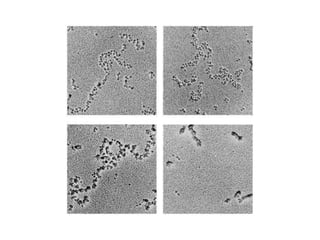
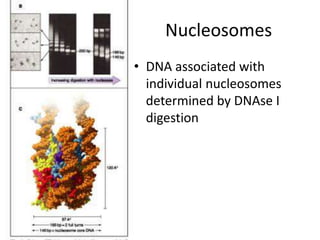

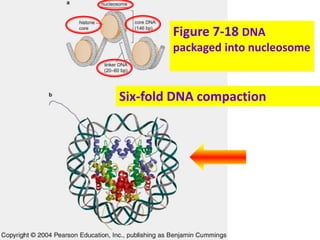

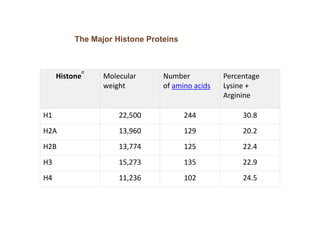
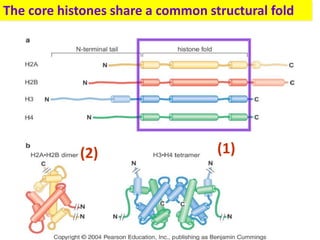
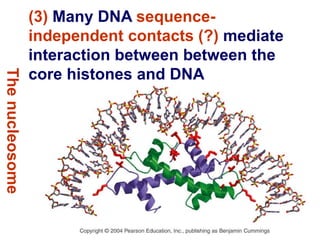
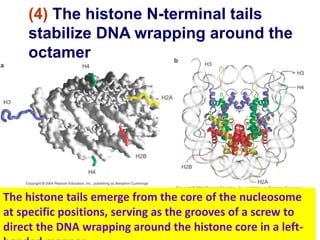
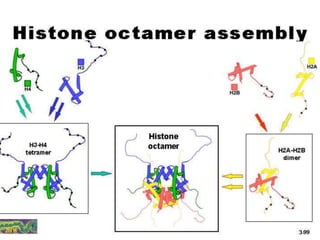
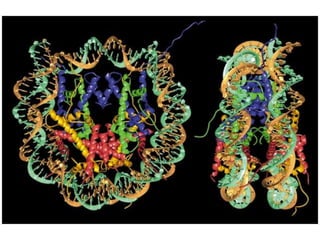
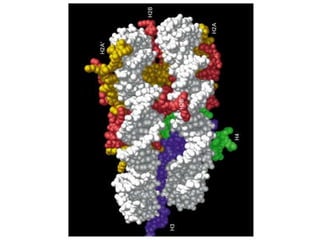










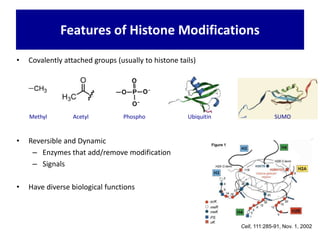
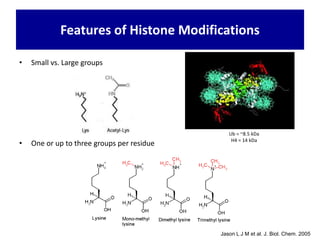
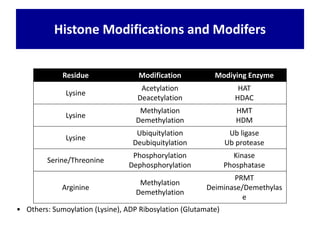



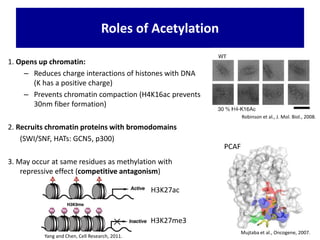
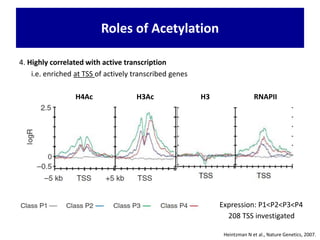
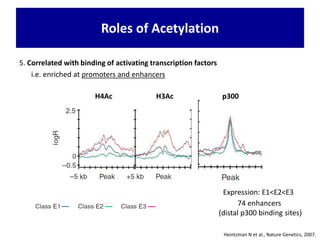

1. The document discusses the structure of chromatin and chromosomes. It describes how DNA is packaged into nucleosomes, which are composed of histone proteins wrapped around DNA. 2. Nucleosomes further compact the DNA to form a 30nm fiber, which is then folded into loops and domains to achieve higher order compaction into chromosomes. This compaction allows the long DNA molecules to fit inside cells. 3. Chromatin structure influences gene expression, with tightly packed heterochromatin generally transcriptionally inactive and loosely packed euchromatin more active. Histone modifications also impact chromatin structure and gene expression.

































































































