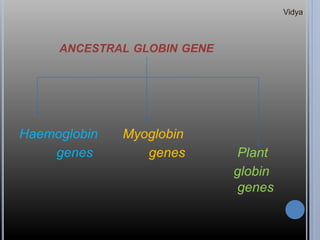
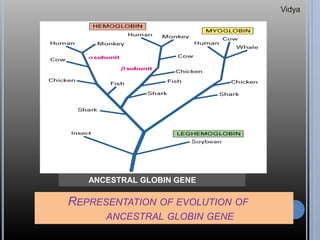
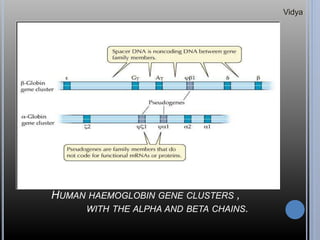
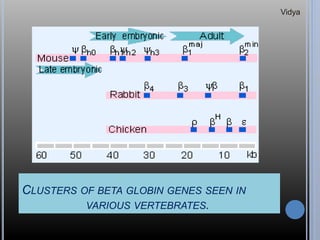
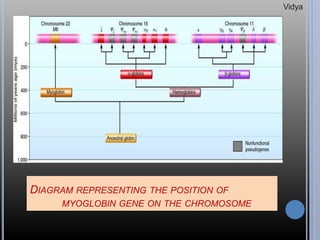
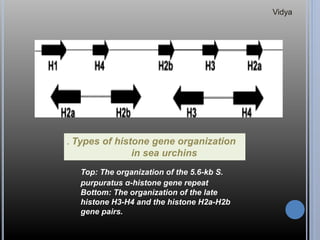
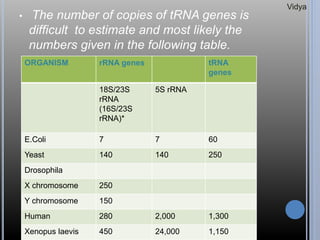
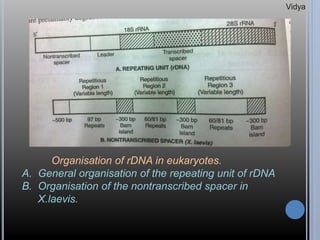
Gene families are sets of similar genes formed by duplication of an original gene. A gene cluster is a subgroup of a gene family where the genes are located near each other on a chromosome. Examples discussed include haemoglobin gene clusters, histone gene clusters, and ribosomal RNA gene clusters. Haemoglobin genes are expressed at different developmental stages. Myoglobin is related to haemoglobin and encodes oxygen transport in muscle. Histone genes encode structural proteins that package DNA into nucleosomes. Ribosomal RNA genes are present in high copy numbers and encode components of ribosomes.