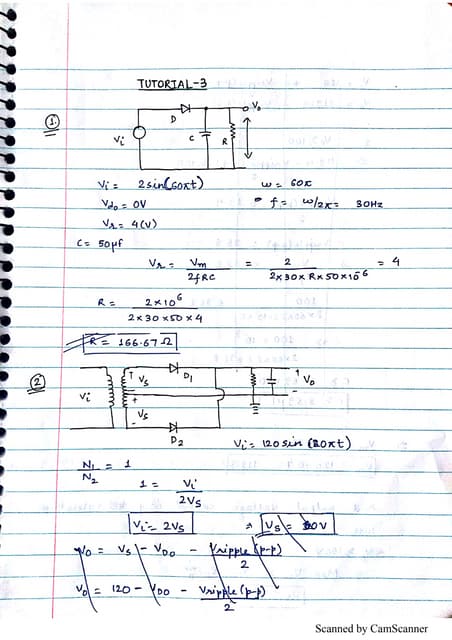
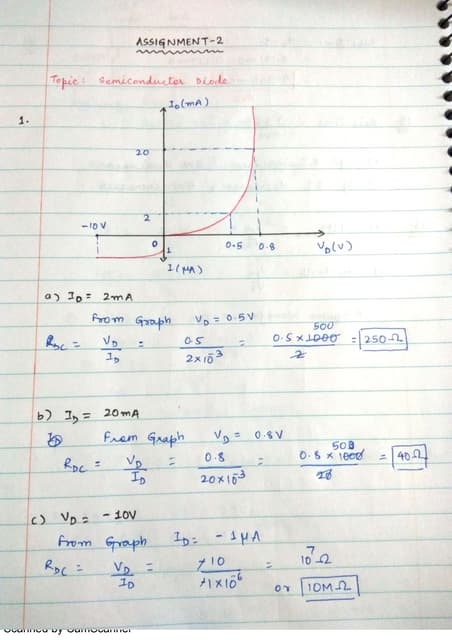
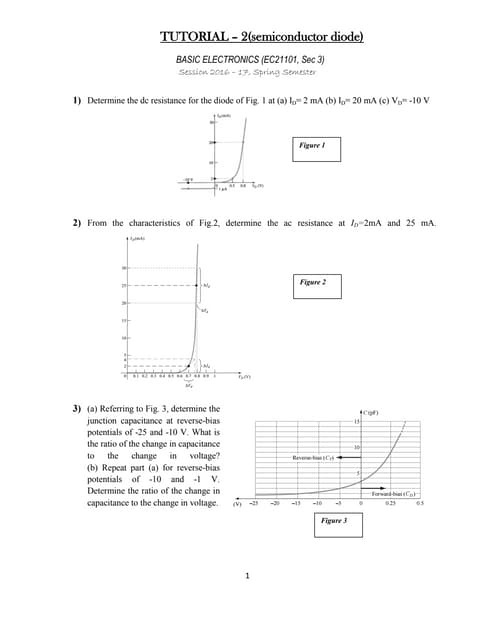
Downloaded 220 times

































![
ES
SE
1
1
k
k
KD
KD is the dissociation constant for the ES complex.
Formation of enzyme-substrate complex
E S
k1
k1
ES
k2
E P
At equilibrium:
forwards reaction rate = reverse reaction rate
i.e. k1[E][S] = k-1[ES]](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-34-320.jpg)

![ ESEE T 3
Combining 1 + 2 + 3
ESkkSES-Ek 21-T1
SEkSkkkES T1121-
rearranging
Divide by k1 and solve for [ES] Where
1
21-
k
kk
K
M
SK
SE
ES T
M](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-36-320.jpg)


![low [S], v is proportional to [S] - first order
high [S], v is independent of [S] - zero order
Michaelis – Menten Kinetics](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-39-320.jpg)

![V0 = Vmax [S]
Km + [S]
Lineweaver-Burk plot transforms the Michaelis-
Menten equation into linear form.
1 = Km + [S]
V0 Vmax [S]Lineweaver-Burk Plot
The double reciprocal plot
Km 1 1
Vmax [S] Vmax
= +
1
V0
(y = mx + c)](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-41-320.jpg)
![For small errors in at low [S]
leads to large errors in 1/vo](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-42-320.jpg)
![For Michaelis - Menton kinetics k2= kcat
When [S] << KM very little ES is formed and [E] = [E]T
and
SE
K
k
SE
K
k
M
cat
T
M
2
ov
kcat/KM is a measure of catalytic efficiency
V0 = Vmax [S]
KM + [S] kcatk 2
T
max
E
V
The turnover number (kcat)
kcat is how many reactions an enzyme can catalyze per second](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-43-320.jpg)
![KM
Relates to how strongly an enzyme binds its substrate.
High KM means strength of binding is low.
kcat
Relates to how rapid a catalyst the enzyme is.
High kcat means high speed of catalysis.
Vmax
Related to kcat and [ET] by: Vmax=kcat[ET]
High Vmax means high rate of catalysis.](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-44-320.jpg)
![A high kcat/KM ratio implies an efficient enzyme
This could result from: Large kcat
Small KM
• kcat = turnover number; kcat = Vmax/[ET]
• kcat/Km is a measure of activity, catalytic efficiency
KM is a useful indicator of the affinity of an enzyme
for the substrate
A low KM indicates a high affinity for the substrate](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-45-320.jpg)



![Competitive Inhibition
(y = mx + c)
Km 1 1
Vmax [S] Vmax
= +
1
V0](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-49-320.jpg)
![Competitive Inhibition
(y = mx + c)
Km 1 1
Vmax [S] Vmax
= +
1
V0
α = 1 + [I]/Ki](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-50-320.jpg)

![Uncompetitive Inhibition
Uncompetitive inhibitors bind at a site distinct from the substrate active site
and bind only to the ES complex
• Active site distorted after binding of S (usually
occurs in multisubstrate enzymes) Decreases both
KM and kcat
• Vo = Vmax[S]/(KM + ’[S]) K’I = [ES][I]/[ESI]
• Cannot be reversed by increasing [S] – available
enzyme decreases](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-52-320.jpg)
![Uncompetitive Inhibition
(y = mx + c)
α’ = 1 + [I]/K’i
Km 1 1
Vmax [S] Vmax
= +
1
V0](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-53-320.jpg)
![Inhibitor can bind at a site distinct from the substrate active site
to either E or ES.
Mixed (Non-competitive) Inhibition
• Vo
= Vmax[S]/(KM + ’[S])
• Vmax decreases; KM can
go up or down.](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-54-320.jpg)
![Non-competitive Inhibition
(y = mx + c)
Km 1 1
Vmax [S] Vmax
= +
1
V0](https://image.slidesharecdn.com/lecture4enzymecatalysisemailed-170211181259/85/Enzyme-Catalysis-55-320.jpg)












This document contains the syllabus and lecture schedule for a course titled "Science of Living Systems". The course covers topics related to protein structure and function including nucleic acids, transcription and translation, enzymes, photosynthesis, respiration, cellular architecture, and cell division. Lectures are scheduled twice a week and include topics like nucleic acids, transcription and translation, protein structure, enzymes, photosynthesis, respiration, and more. The document also lists required textbooks and provides an overview of protein classification, structure, and enzymatic kinetics including Michaelis-Menten kinetics.