Downloaded 234 times












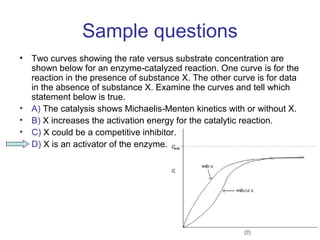
![Vo vs [S] plots give sigmoidal curve
for at least one substrate
Activator can shift hyperbolic (as if there were no T state)
Binding of this allosteric inhibitor or this activator does not
effect Vmax, but does alter Km
Allosteric enzyme do not follow M-M kinetics](https://image.slidesharecdn.com/jftkdotjsskuyedi9hgm-signature-091447bade647bc70325d98b994f7c3a3901d946a34a4516654ab9d79d1d0b85-poli-141026005446-conversion-gate01/85/Lecture-12-16-320.jpg)




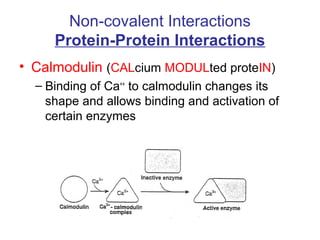
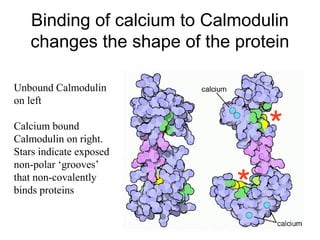
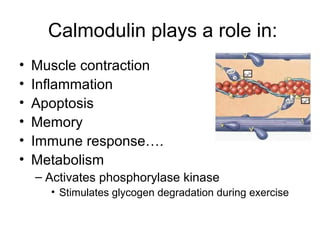
![Calmodulin
• Extracellular [Ca] = 5 mM
• Intracellular [Ca] = 10-4 mM
– Bound Ca can be released by hormonal
action, nerve innervation, light, ….
– Released Ca binds to Calmodulin which
activates a large number of proteins](https://image.slidesharecdn.com/jftkdotjsskuyedi9hgm-signature-091447bade647bc70325d98b994f7c3a3901d946a34a4516654ab9d79d1d0b85-poli-141026005446-conversion-gate01/85/Lecture-12-24-320.jpg)


















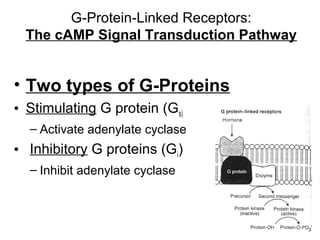
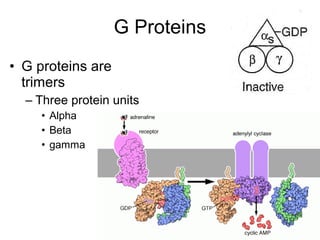

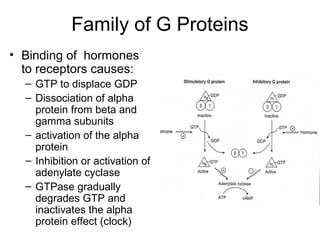
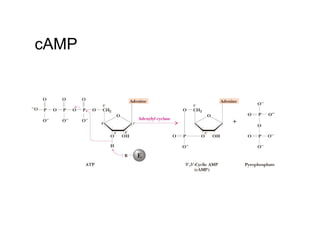
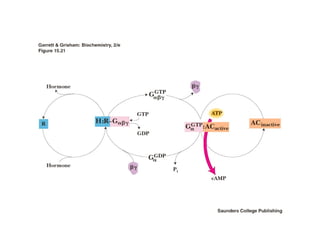
![The cAMP Signal Transduction
Pathway
• cAMP – intracellular messenger
– Elevated cAMP can either activate or inhibit
regulatory enzymes
• cAMP activates glycogen degradation
• cAMP inhibits glycogen synthesis
• [cAMP] affected by rates of synthesis and
degradation
– Synthesis by adenylate cyclase
– Degradation by phosphodiesterase
• Stimulated by insulin
• Inhibited by caffeine](https://image.slidesharecdn.com/jftkdotjsskuyedi9hgm-signature-091447bade647bc70325d98b994f7c3a3901d946a34a4516654ab9d79d1d0b85-poli-141026005446-conversion-gate01/85/Lecture-12-54-320.jpg)
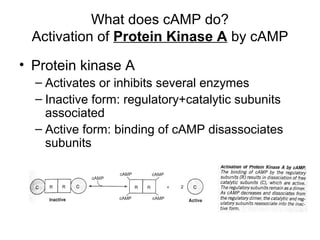

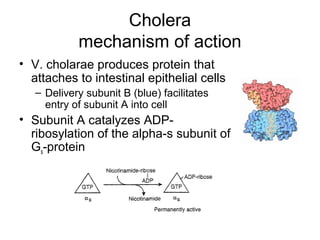







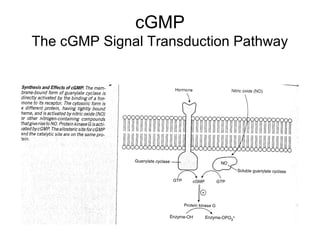

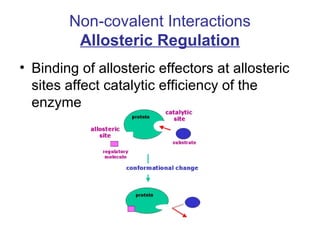
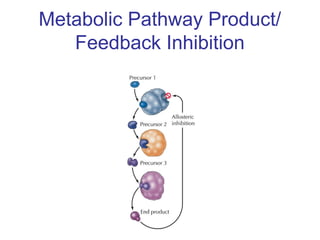
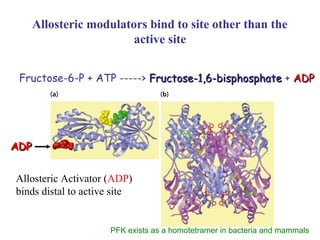
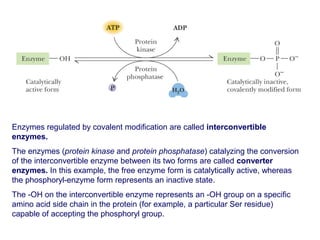
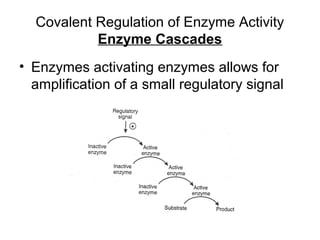
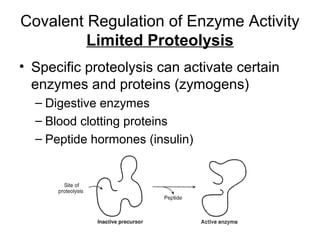
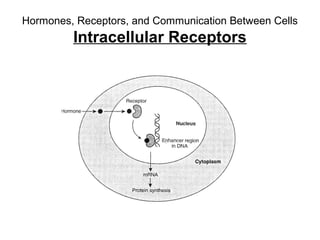
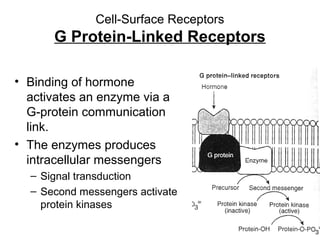
This document discusses various ways that enzyme activity can be regulated. It describes how enzyme levels and activity can be increased or decreased through different mechanisms, including substrate availability, product accumulation, allosteric effectors, covalent modification, genetic controls, zymogens, isozymes, and modulator proteins. A key example discussed is allosteric regulation, where binding of an allosteric effector at a site other than the active site can increase or decrease an enzyme's catalytic efficiency. Phosphorylation is provided as a common example of covalent regulation, where the addition of phosphate groups can alter an enzyme's activity.



















































![16 zoonoses [zoʊ'ɒnəsɪs] pathogens](https://cdn.slidesharecdn.com/ss_thumbnails/16-zoonoseszonsspathogens-150727150950-lva1-app6891-thumbnail.jpg?width=640&height=640&fit=bounds)















