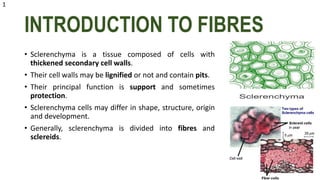
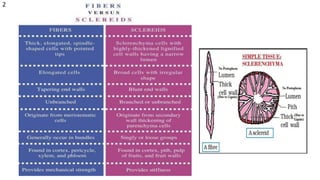
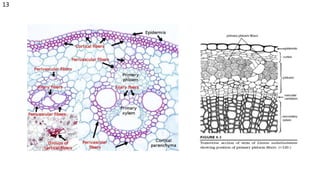
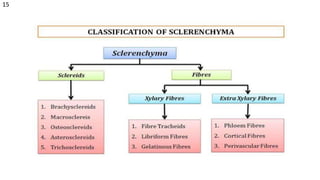
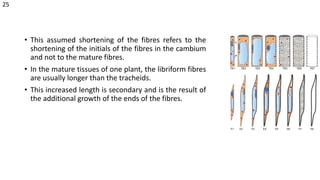
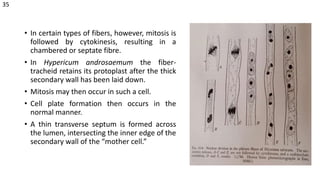
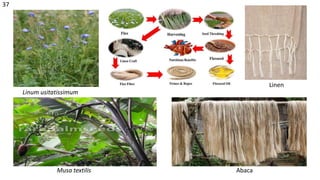
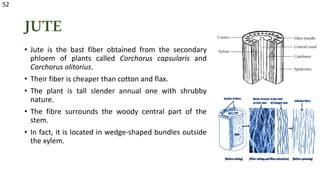
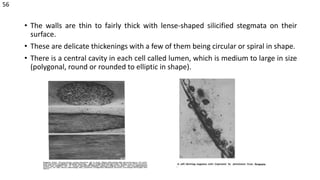
The document details the classification, occurrence, structure, and development of plant fibers, specifically focusing on sclerenchyma and its two primary types: xylary and extraxylary fibers. It describes various types of fibers present in vascular tissues, outlines their evolutionary background, and explains the developmental processes from different meristems. Key distinctions are made between libriform fibers, fiber-tracheids, and their molecular characteristics, emphasizing the functional diversity and structural complexity of these plant fibers.