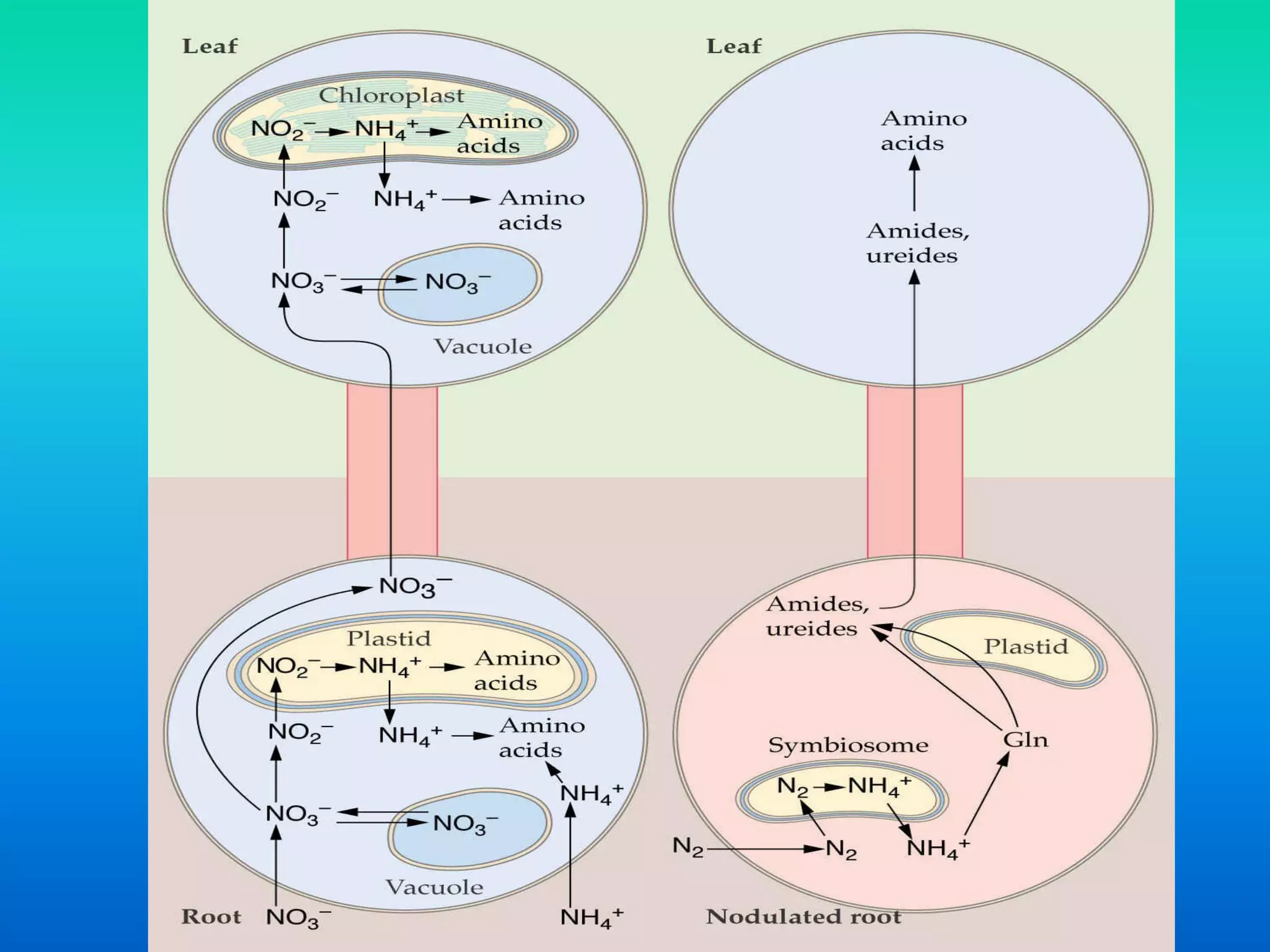
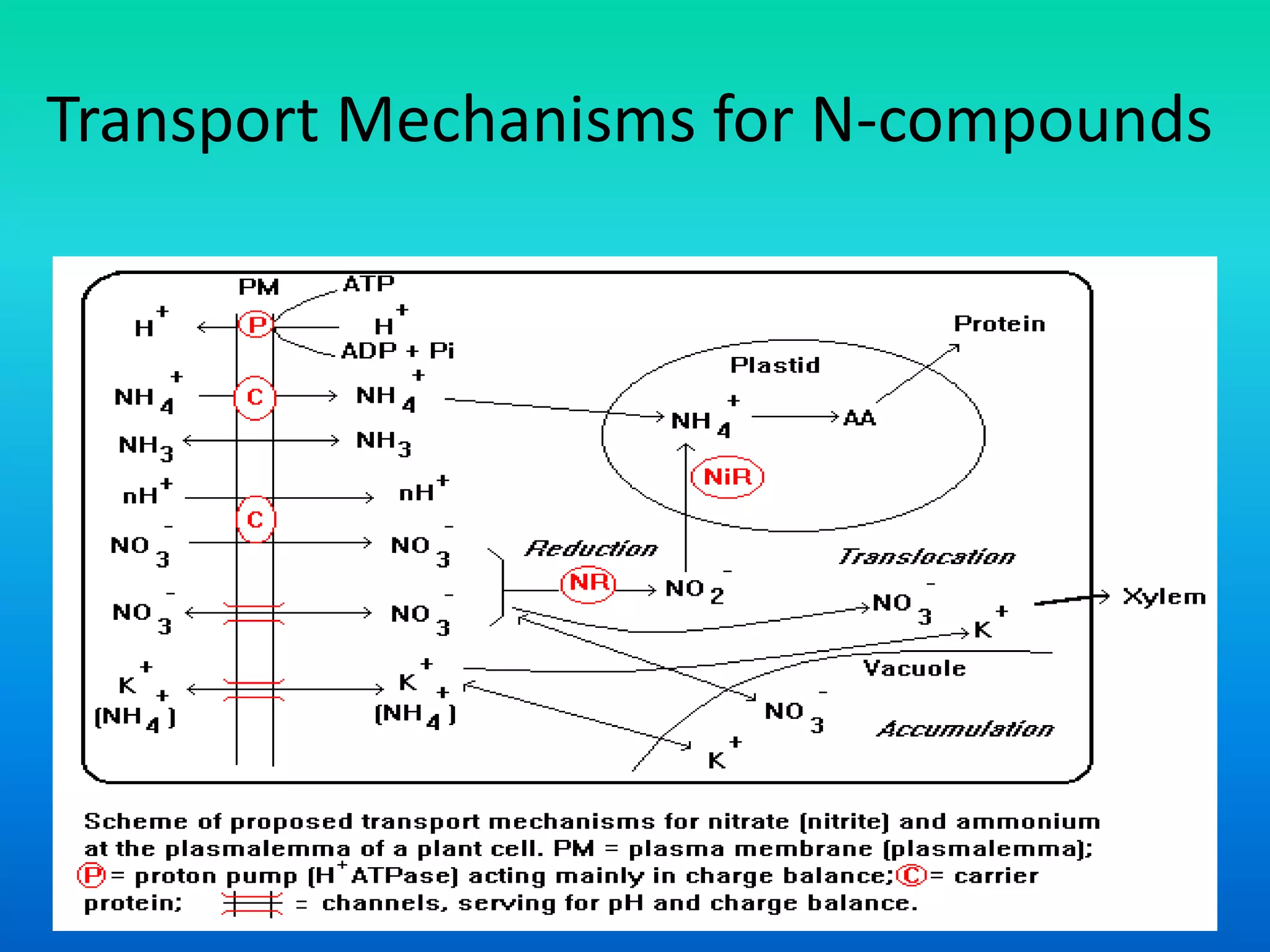
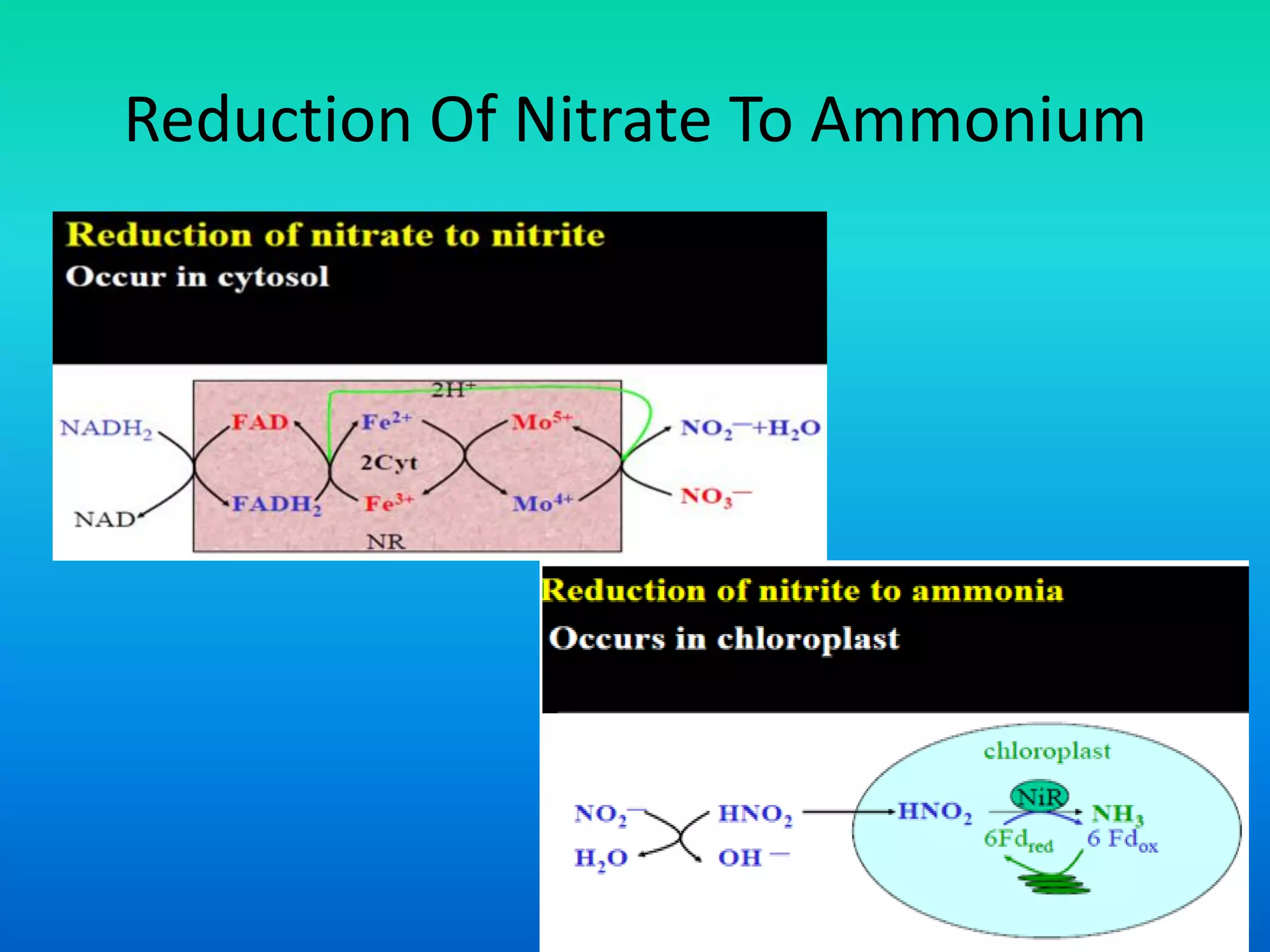
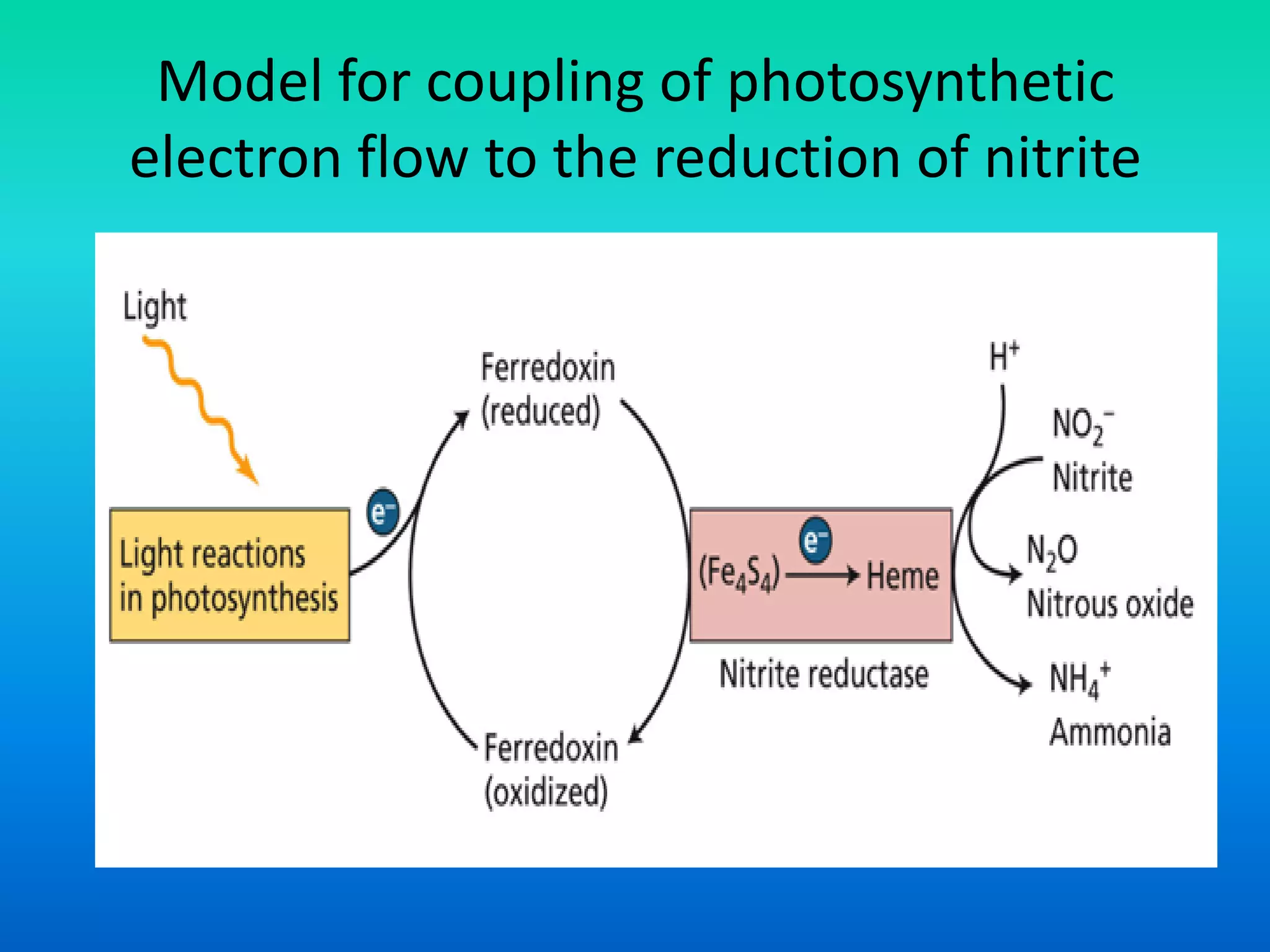
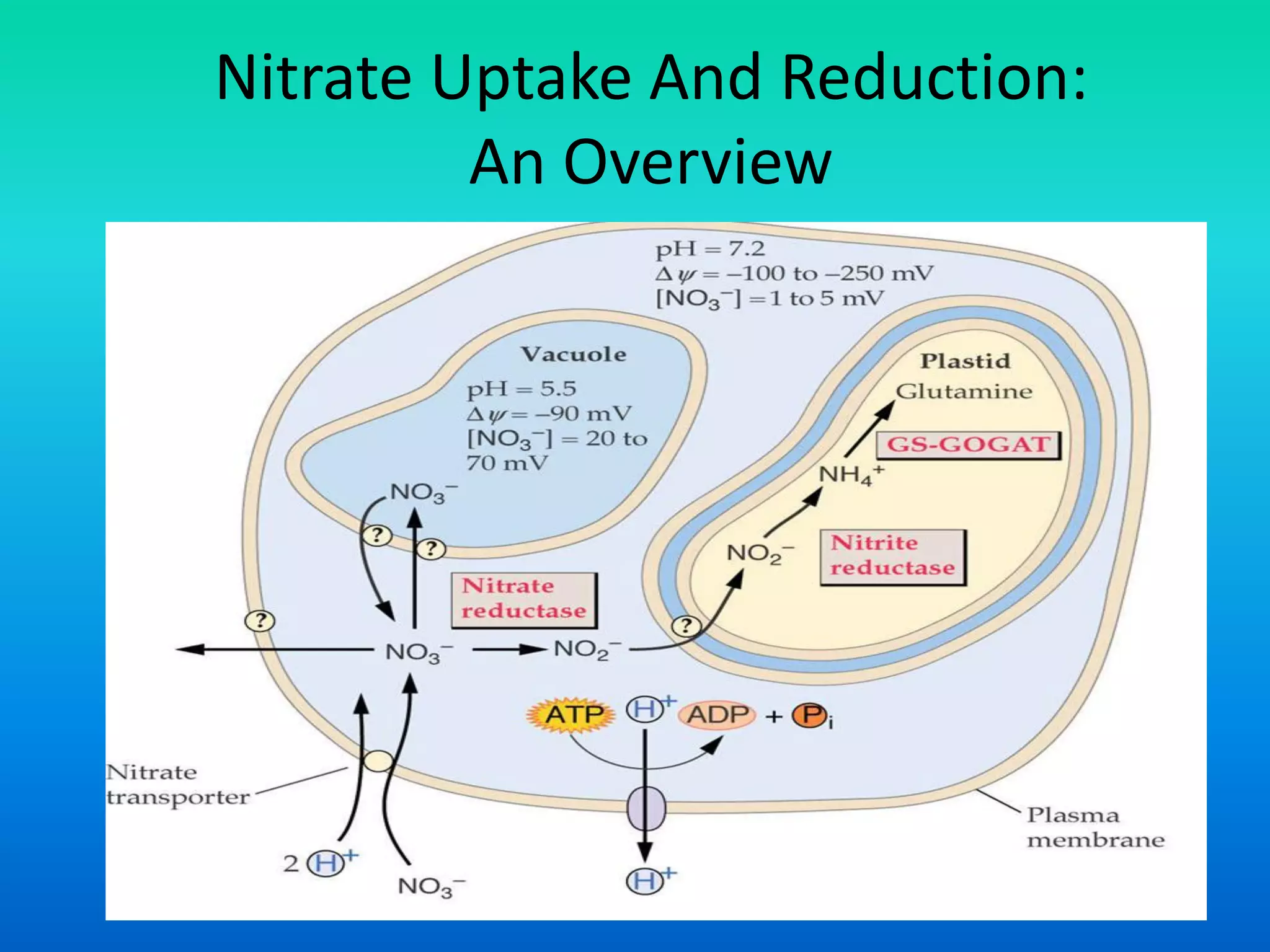
The document discusses the mechanisms of nitrate metabolism in plants, focusing on nitrate uptake, reduction, and assimilation processes. It describes the roles of various transport systems, enzymes like nitrate reductase and nitrite reductase, and the interplay between carbon and nitrogen metabolism. Additionally, it covers regulation of nitrate uptake and the physiological implications of nitrate presence in plants.