Downloaded 180 times
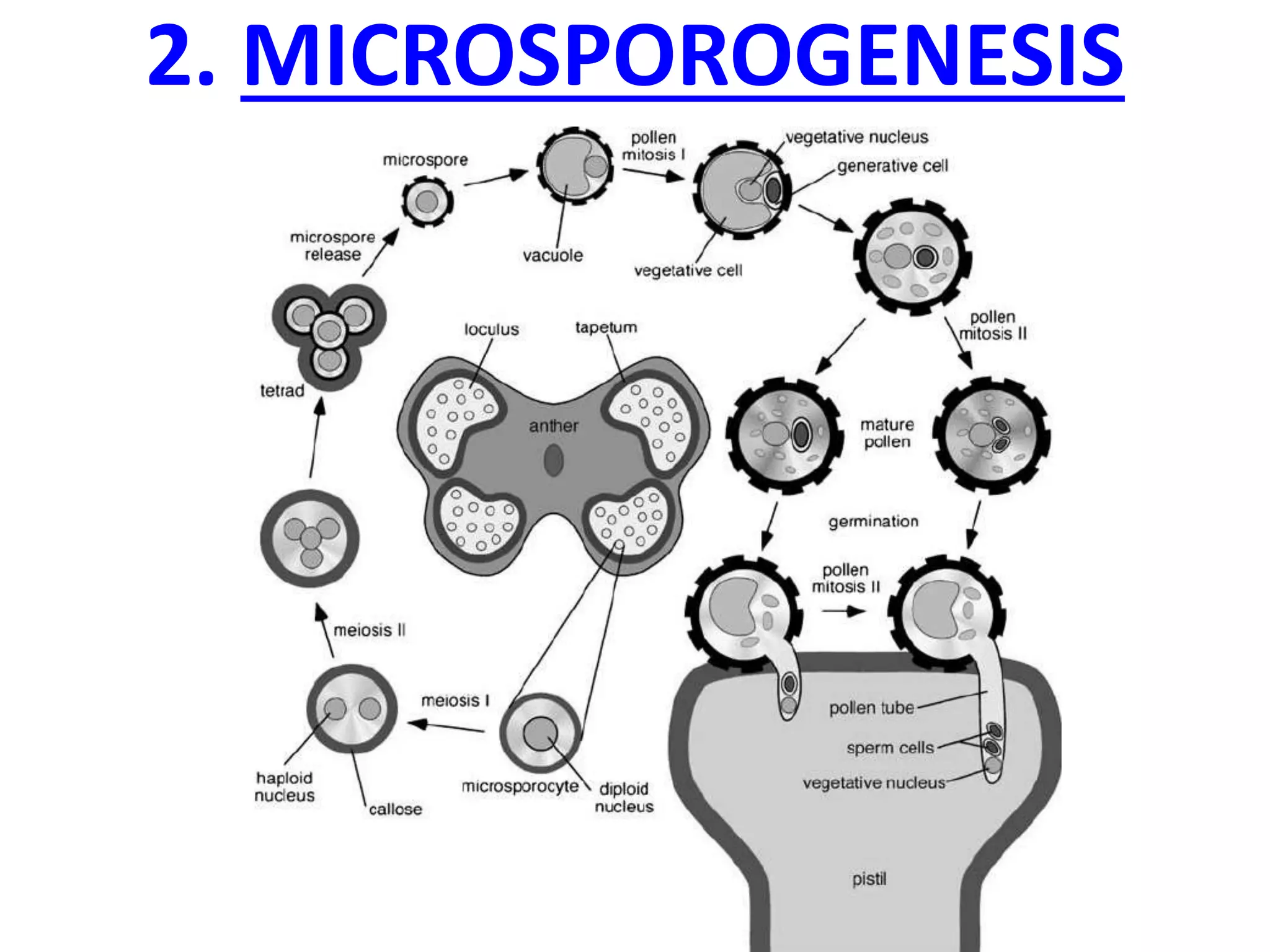

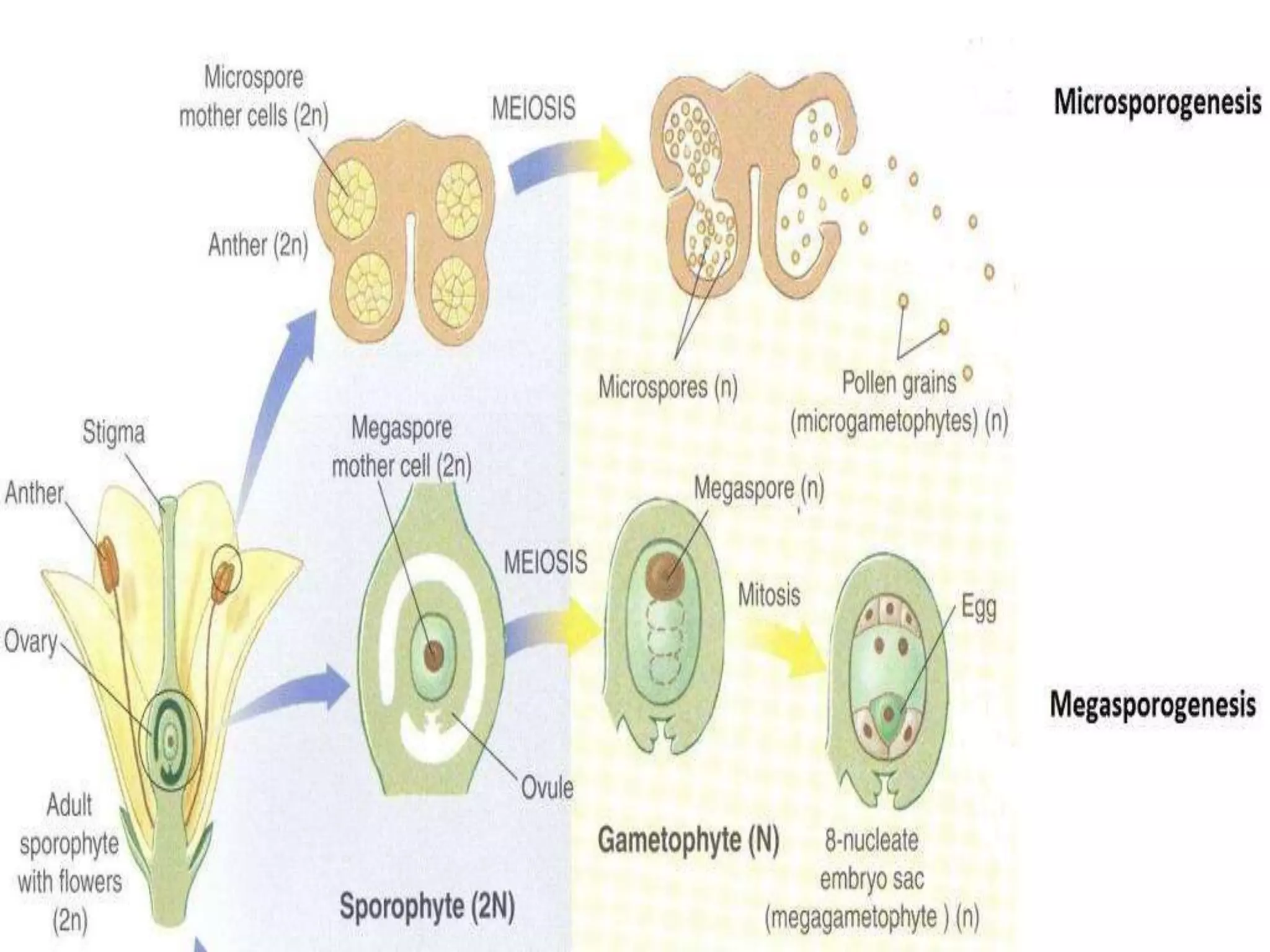



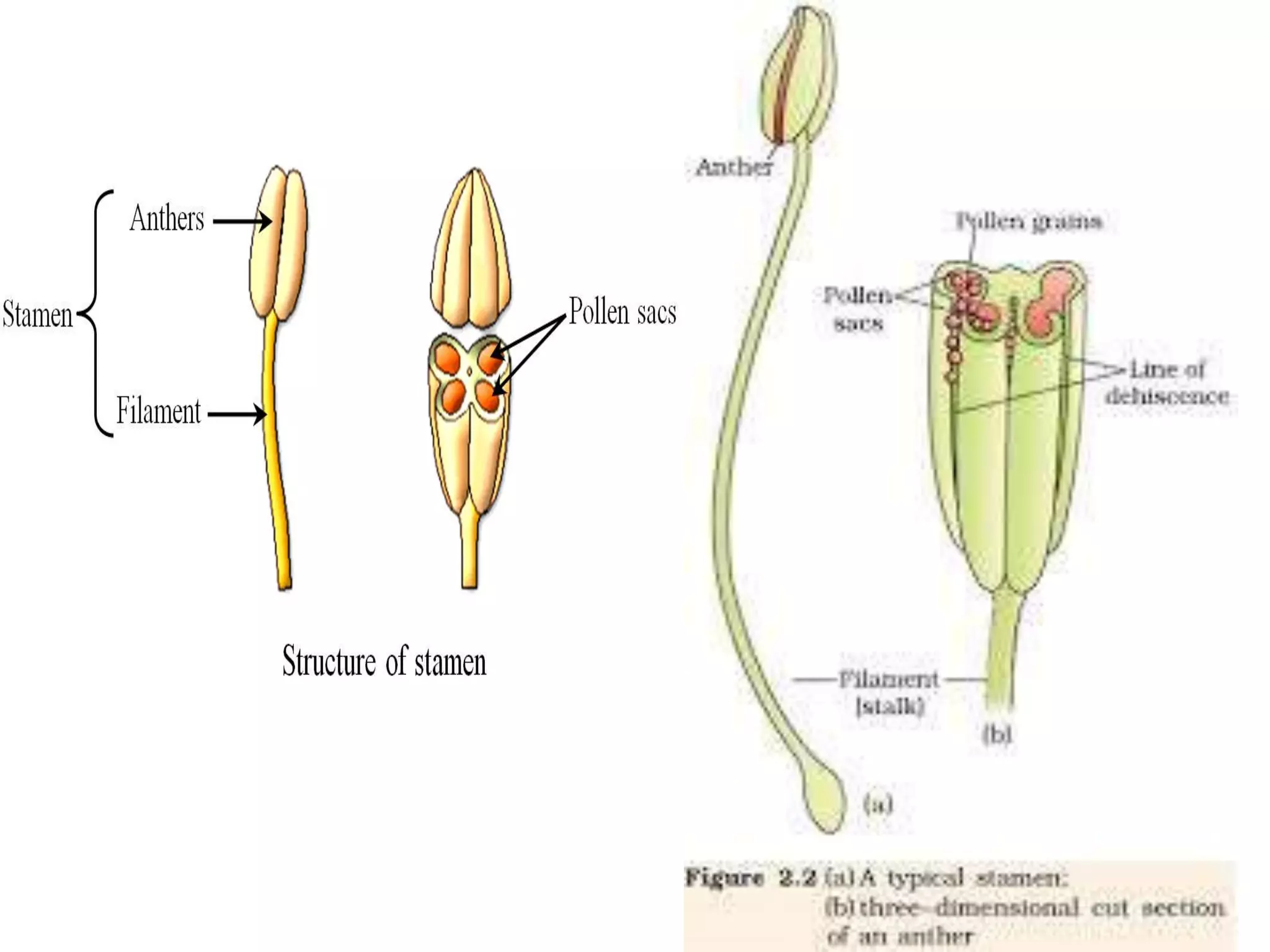
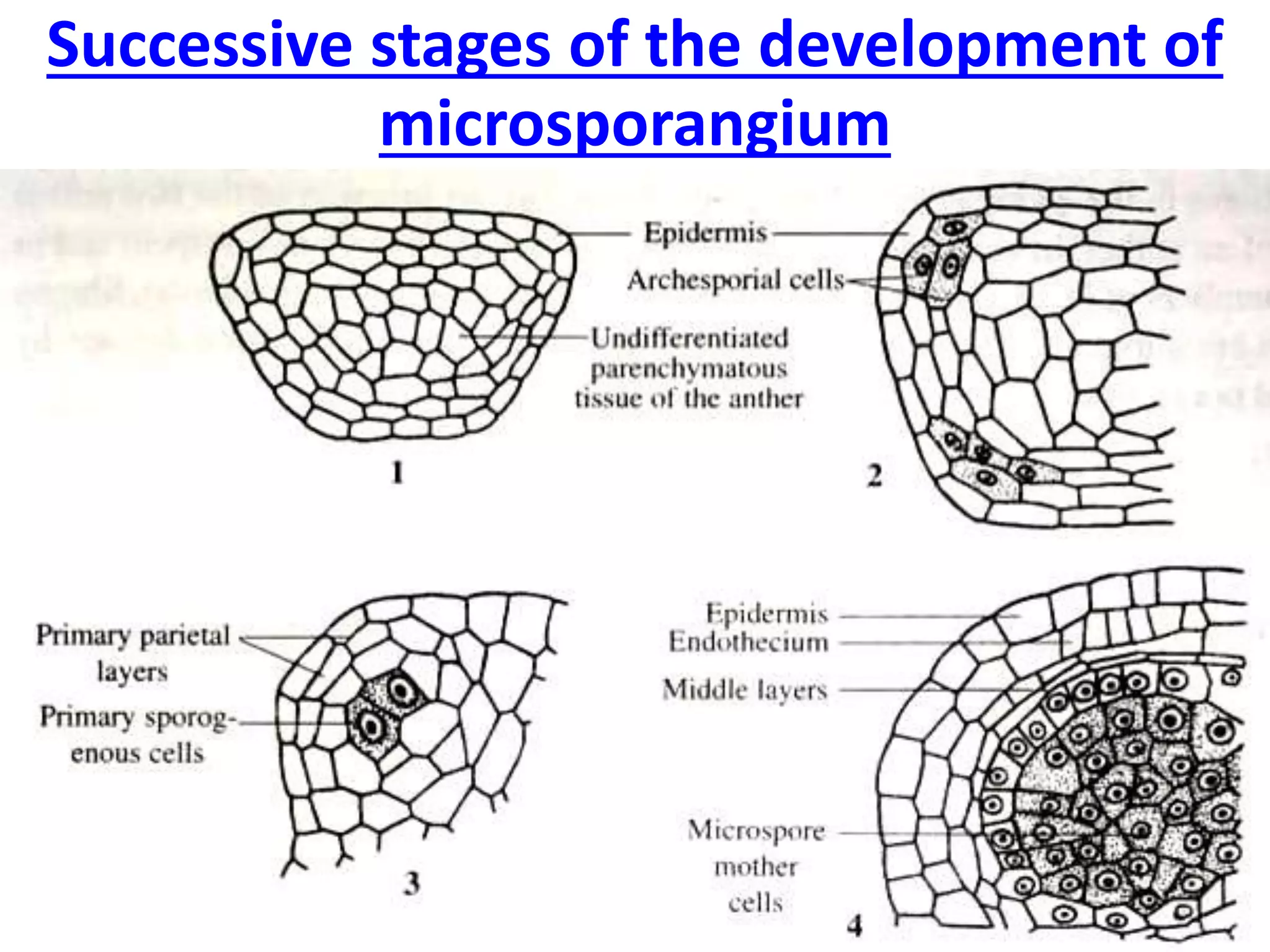
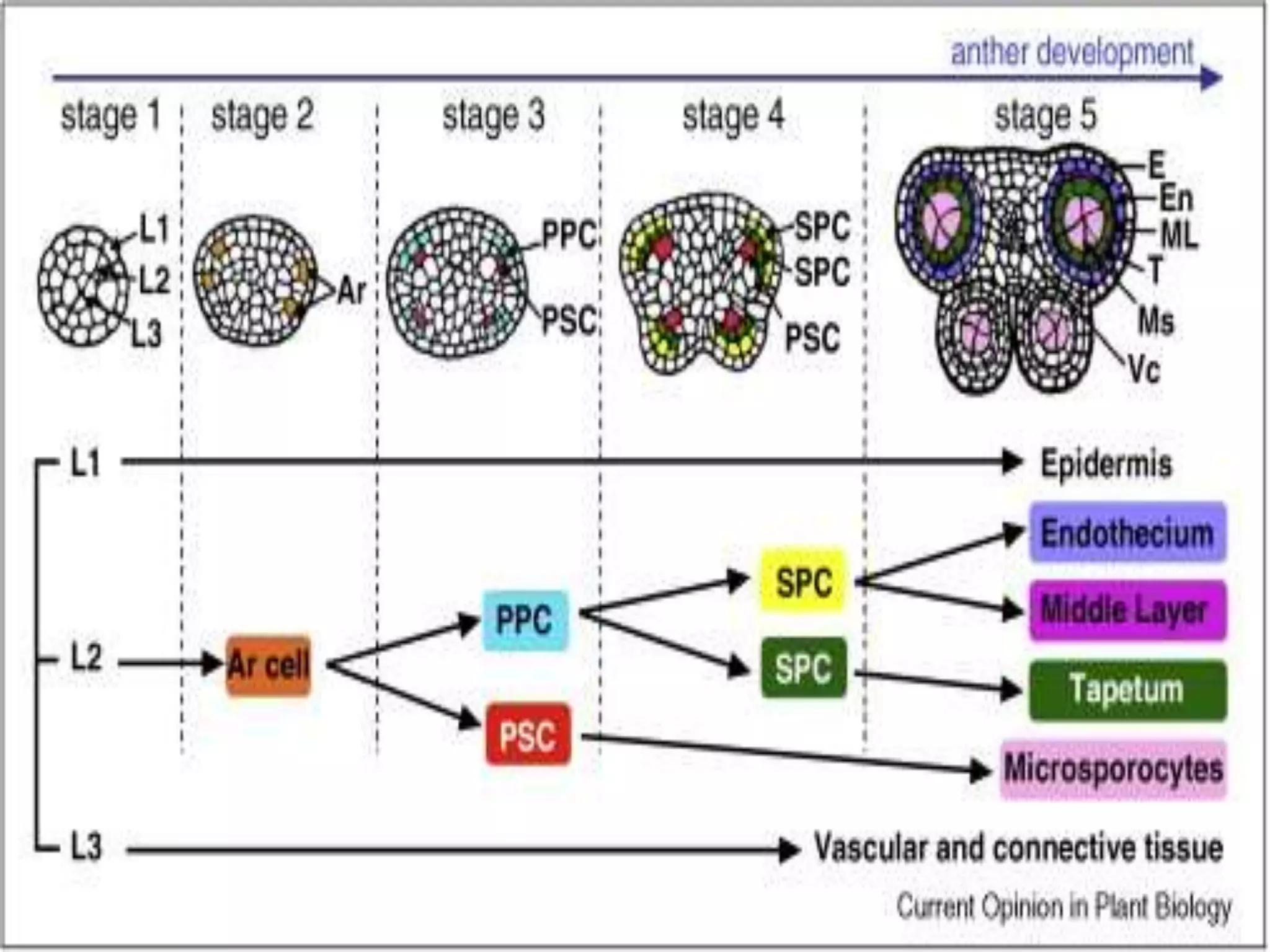

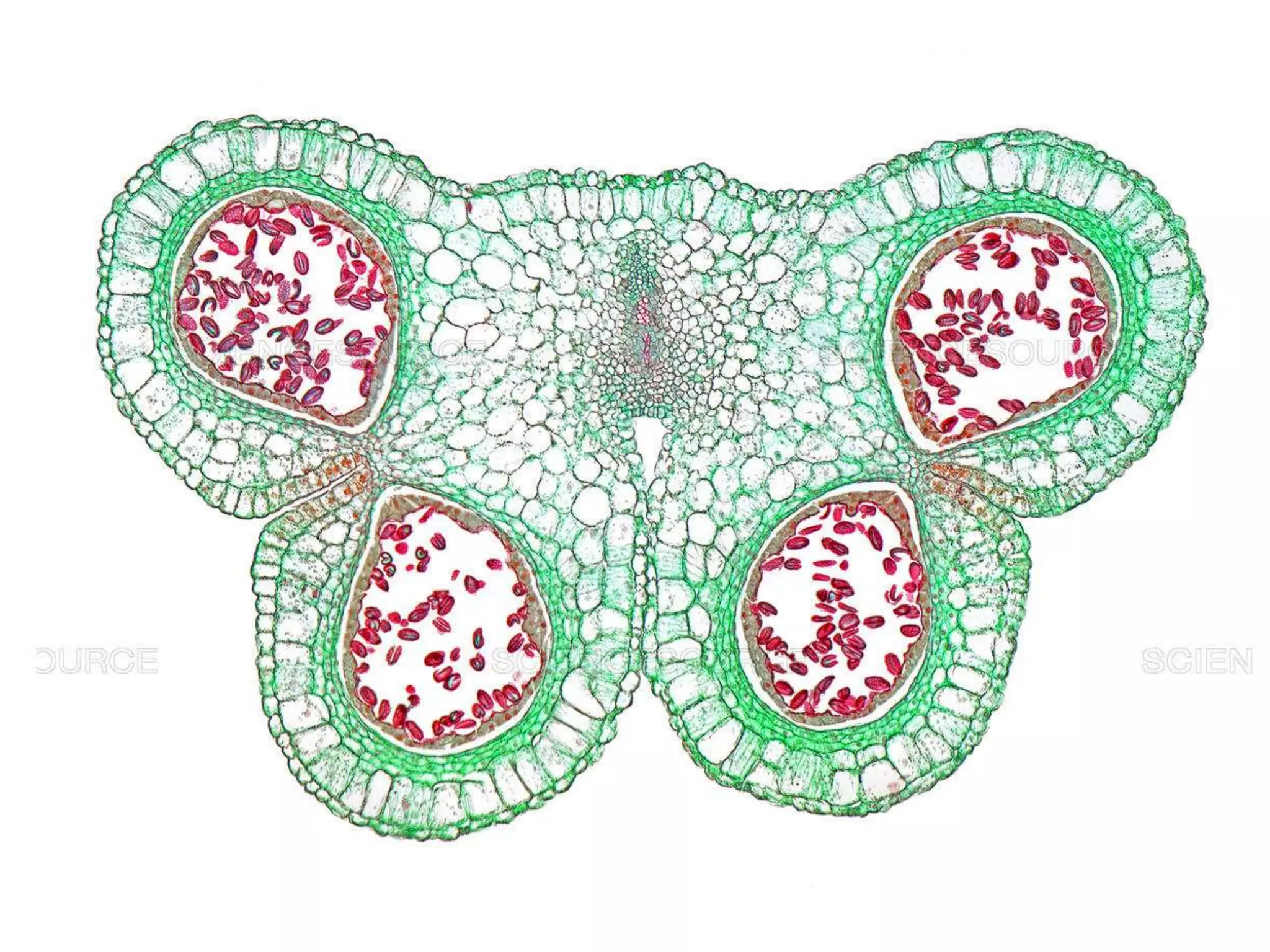




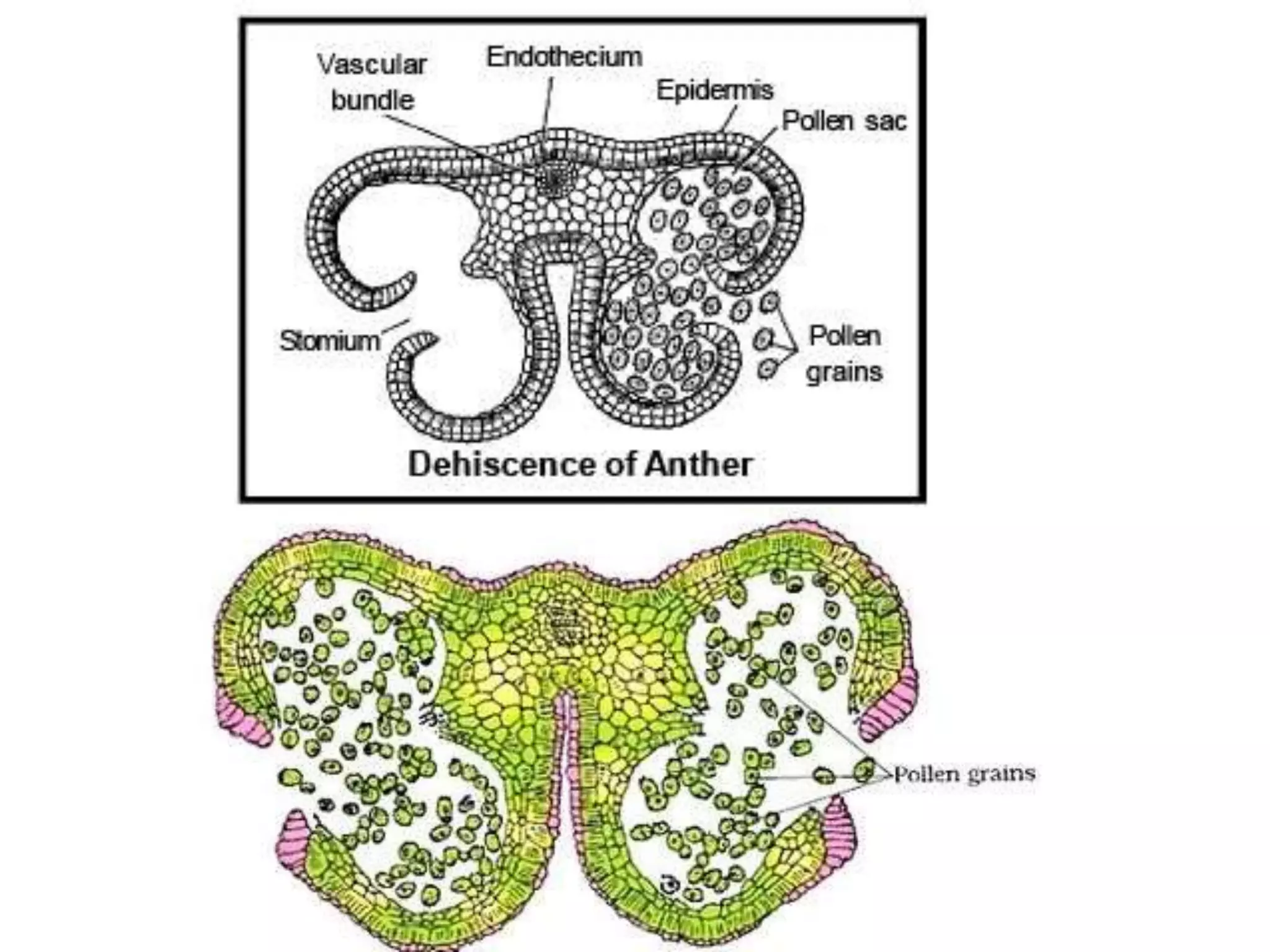

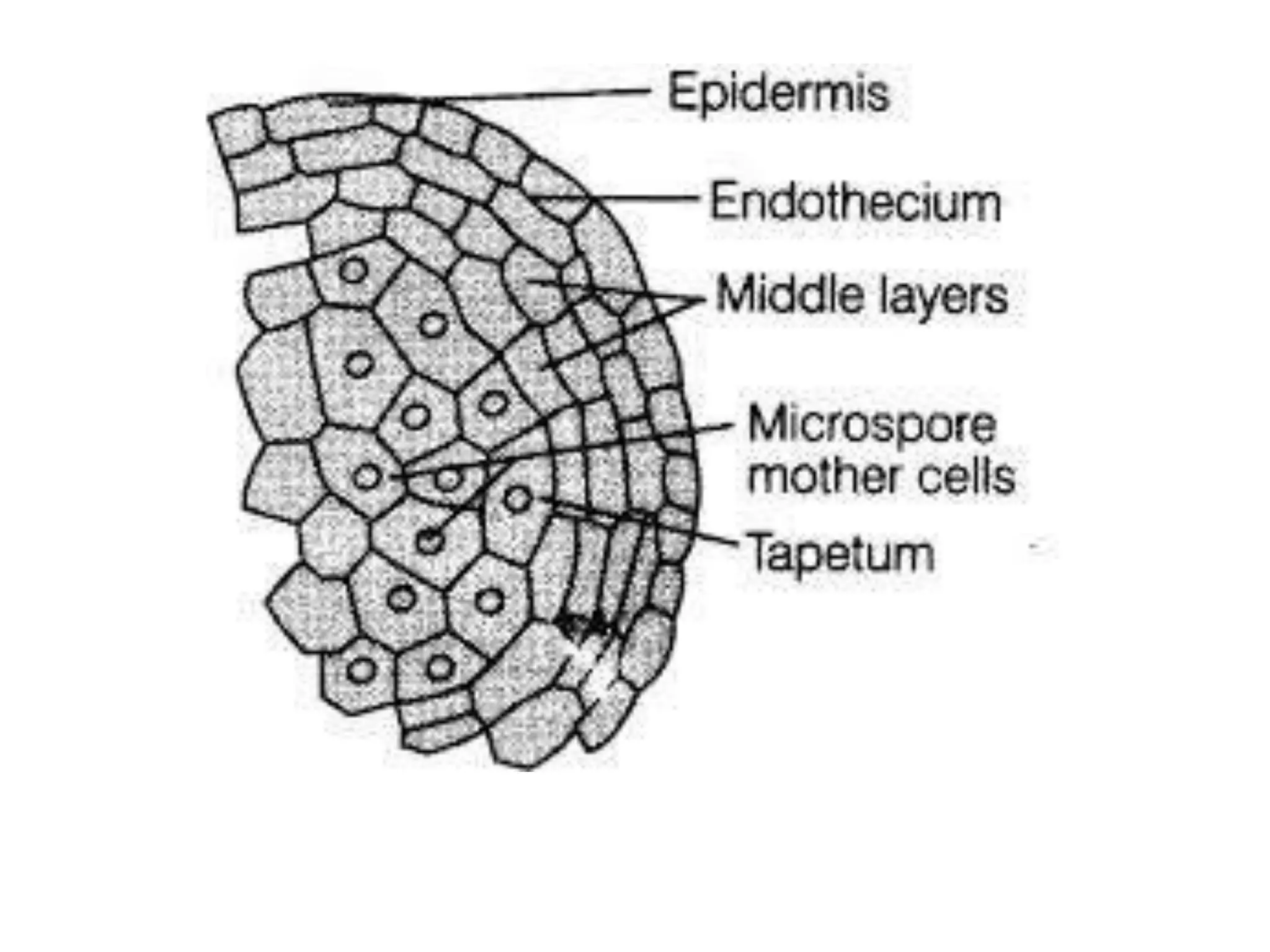



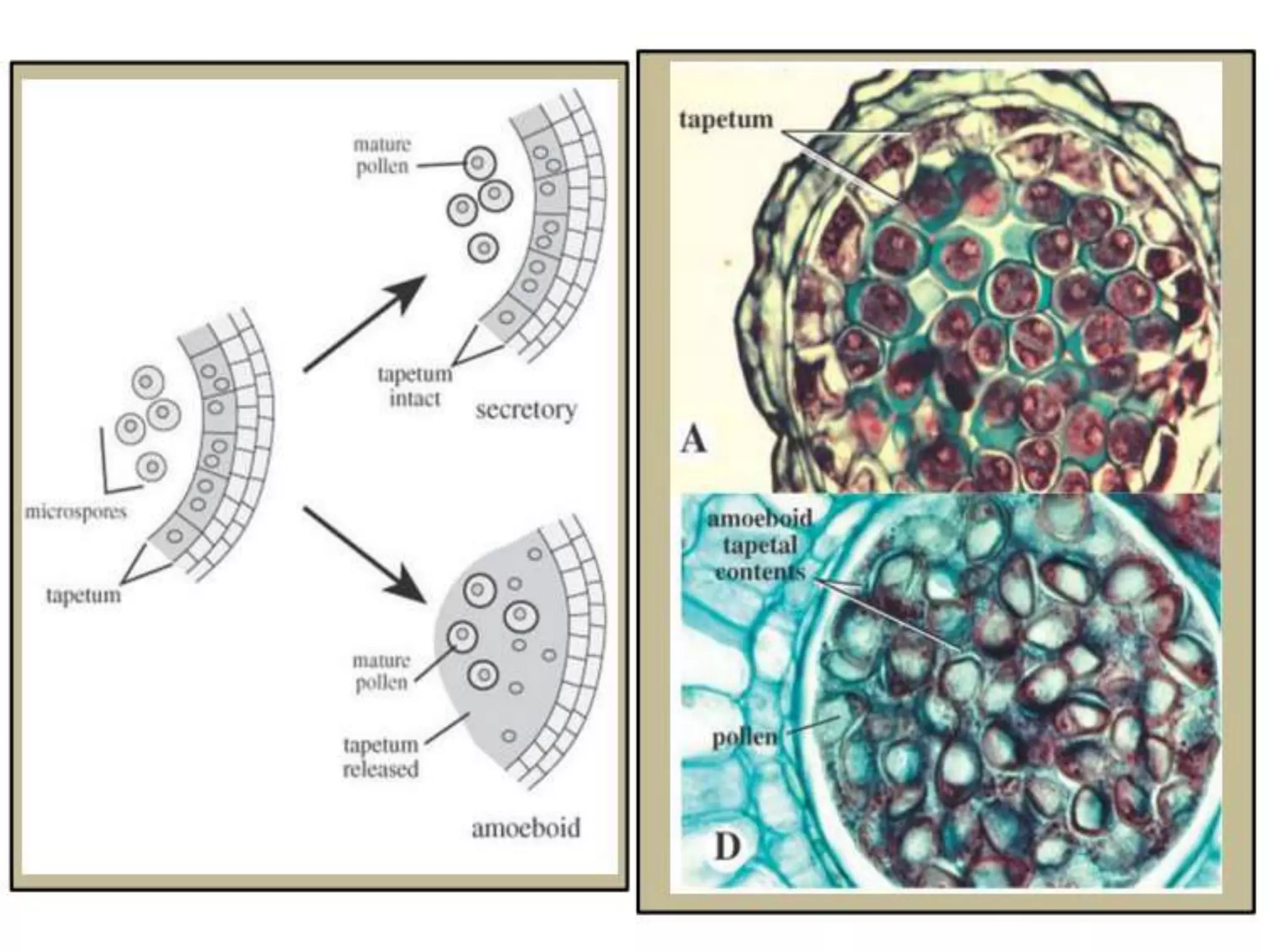






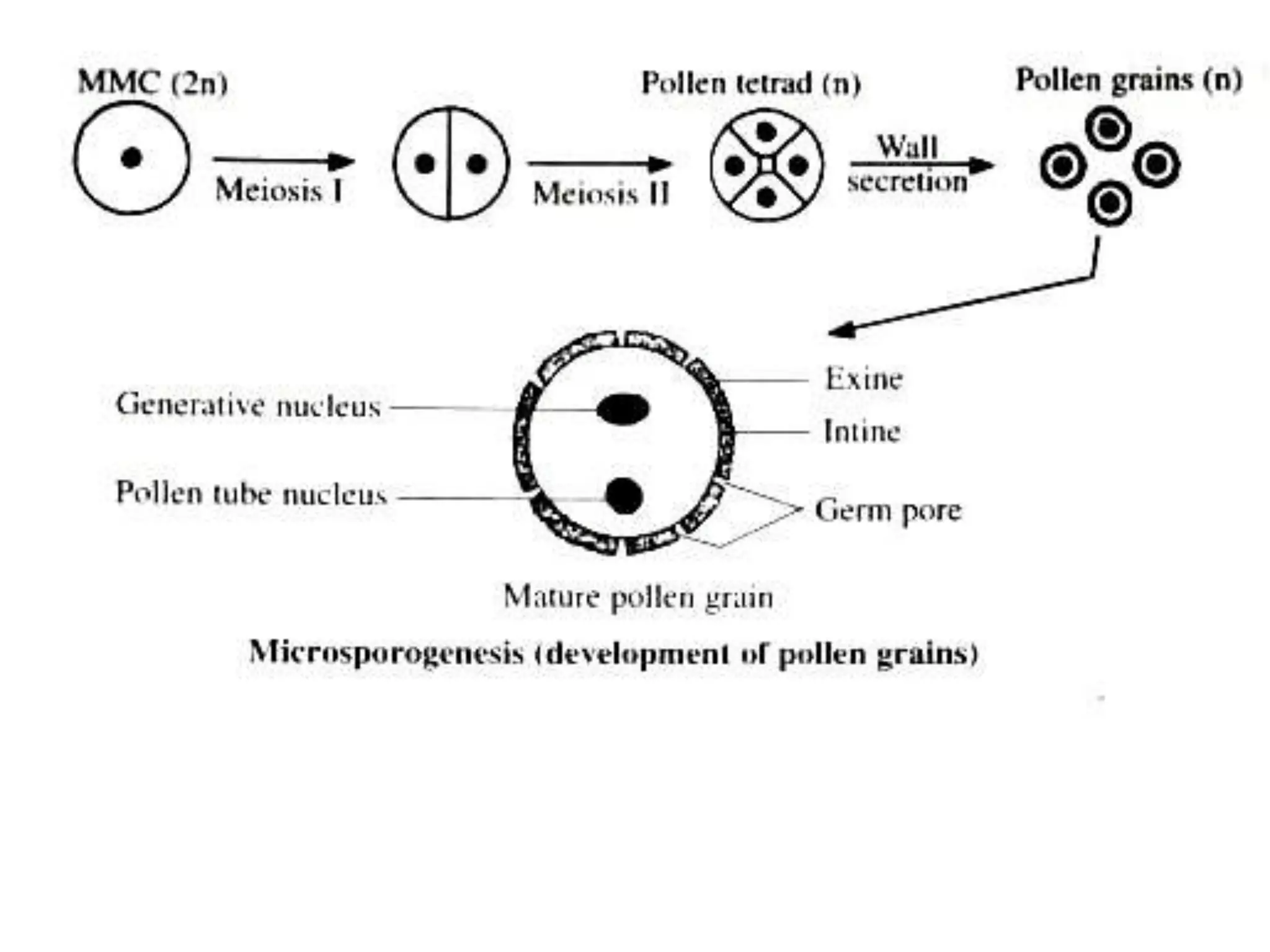
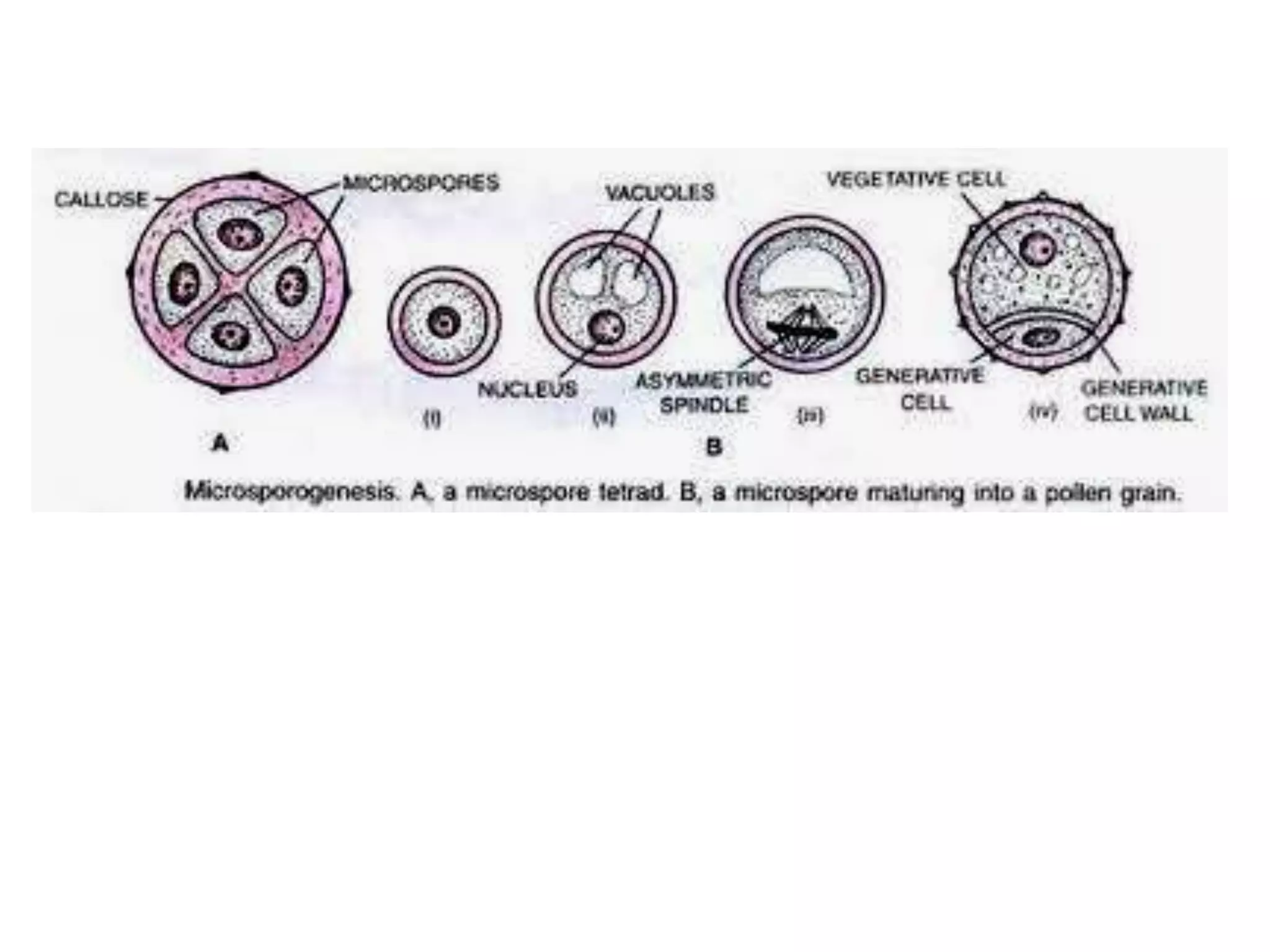


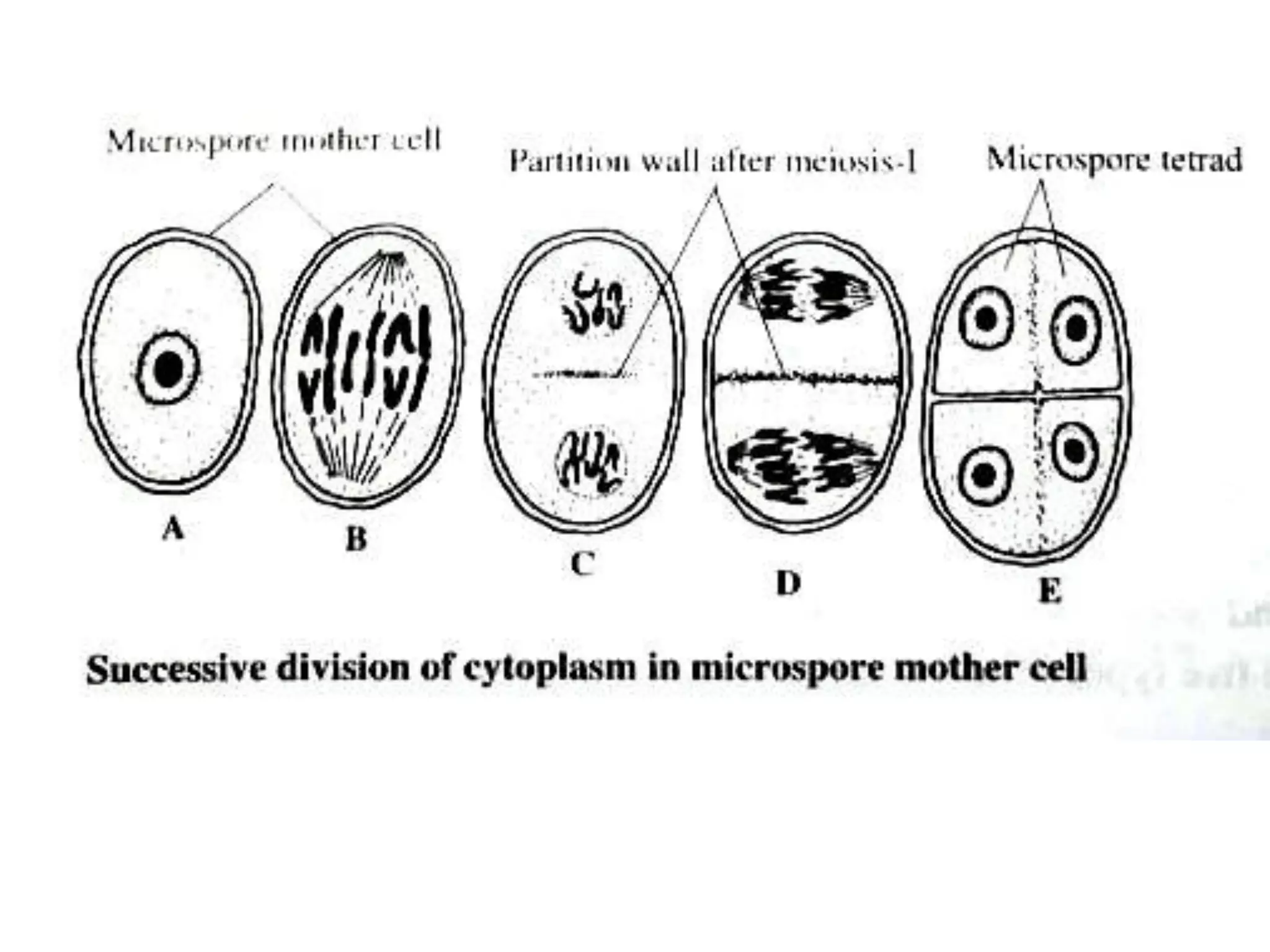

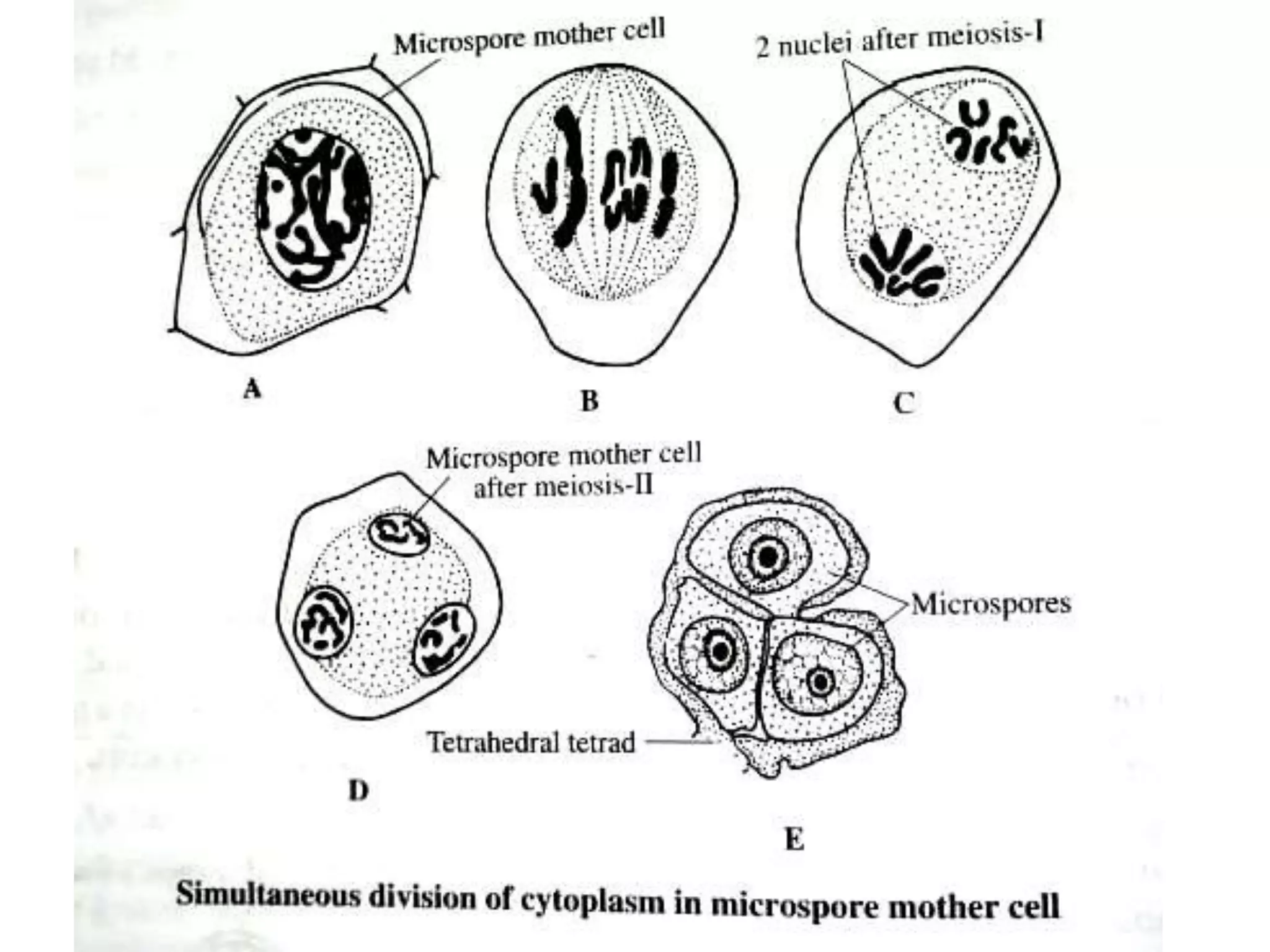

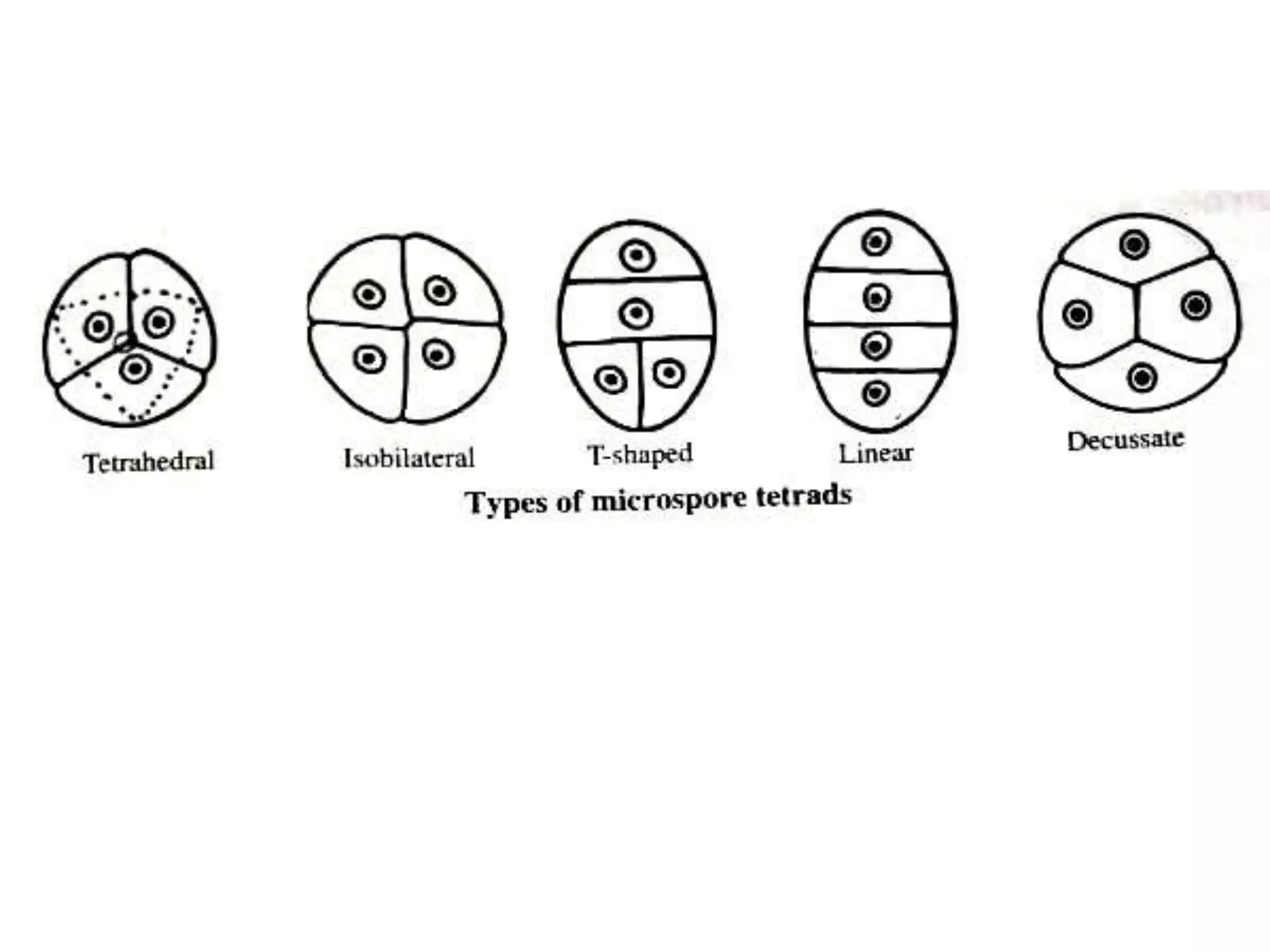











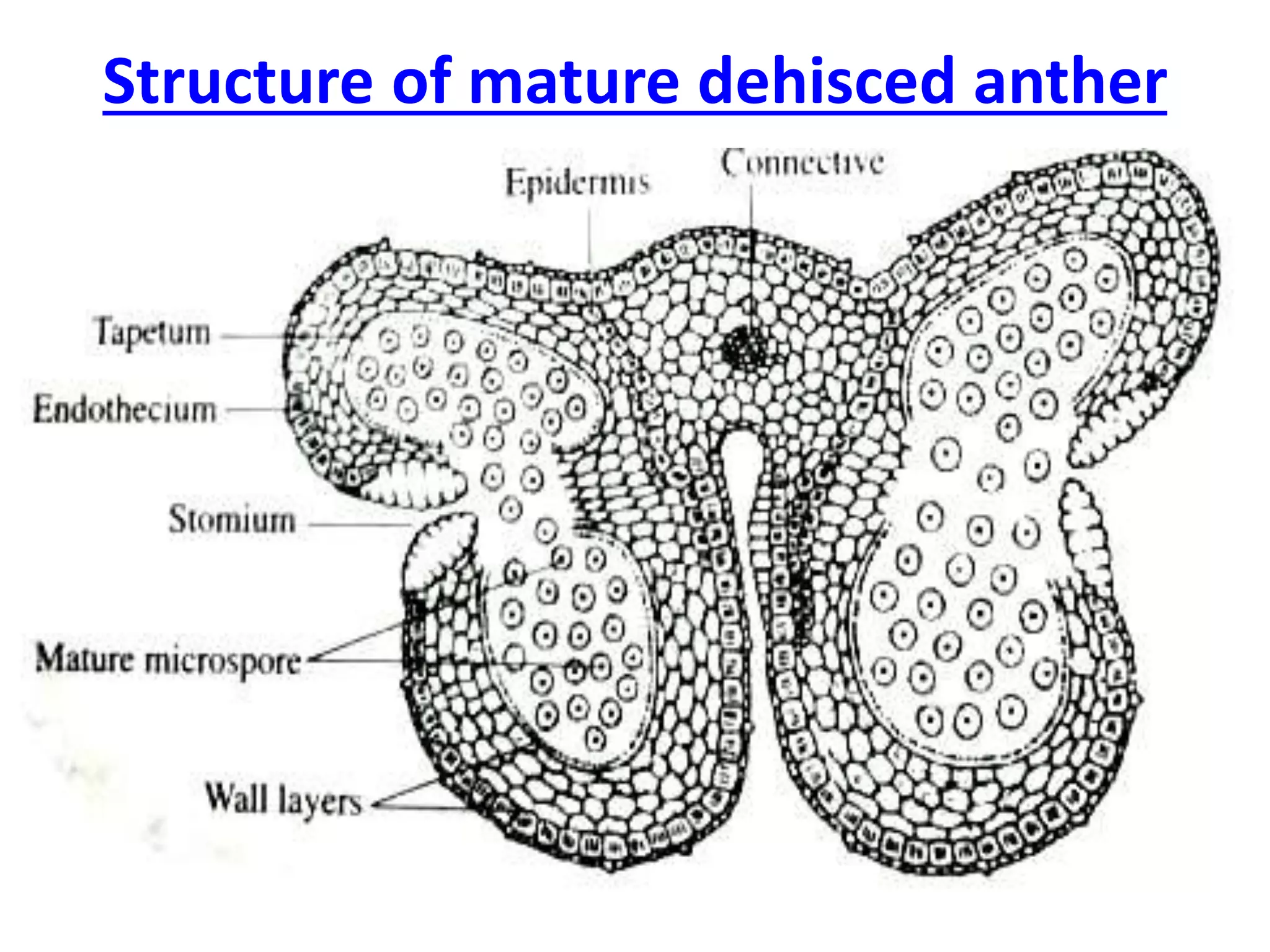







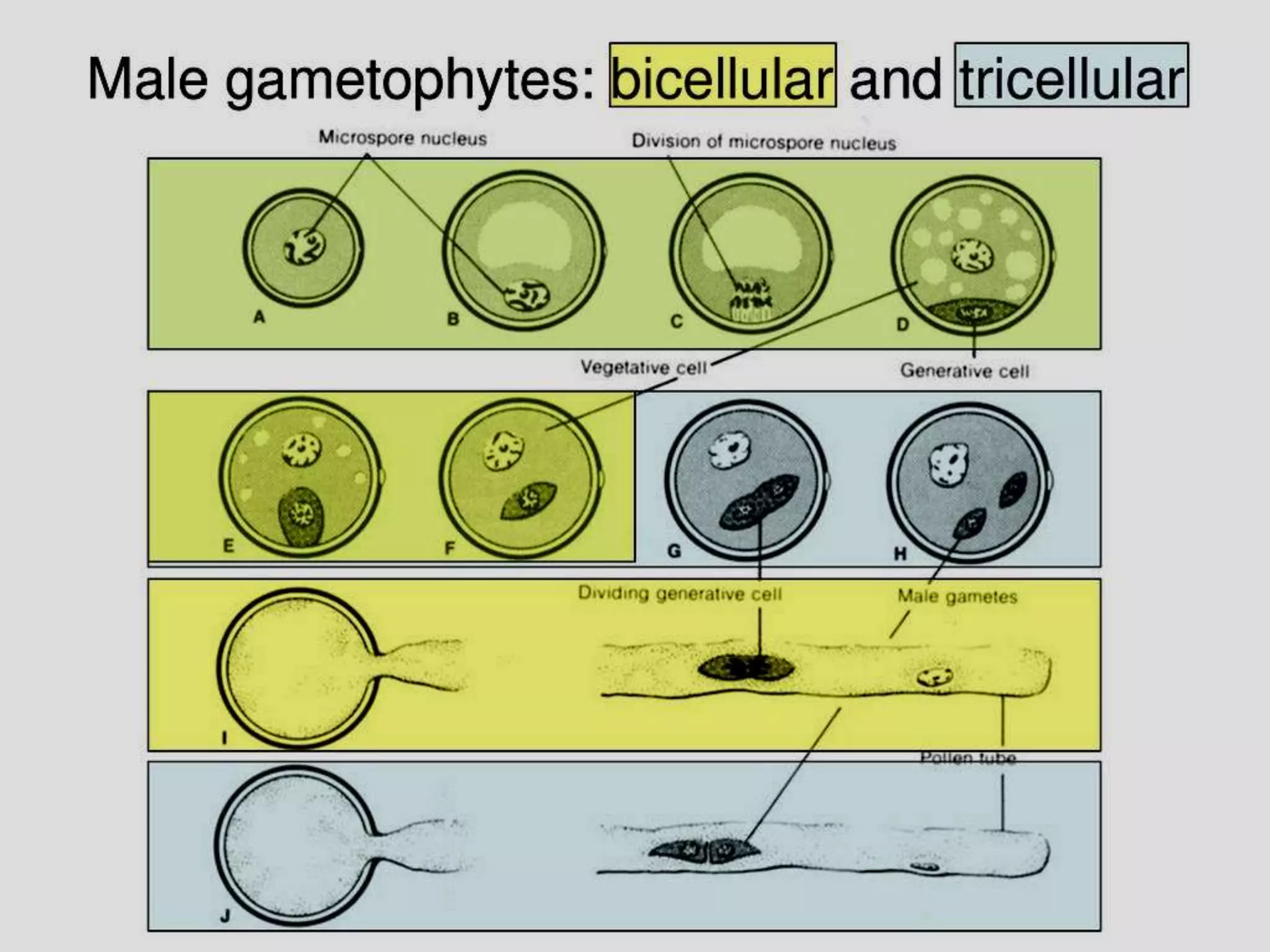






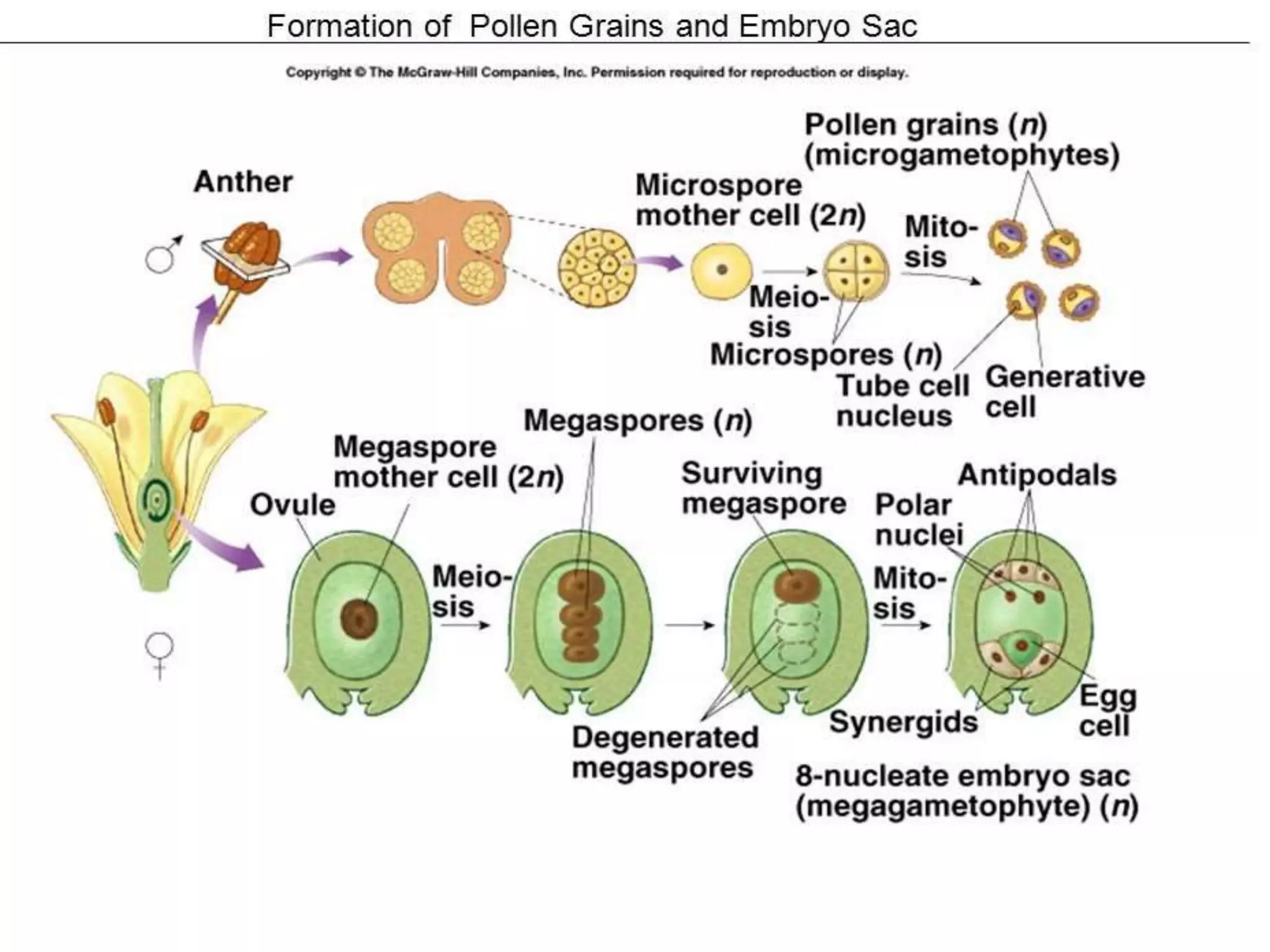




Microsporogenesis involves the formation of pollen grains in the anthers. It begins with the formation of archesporial cells that develop into primary sporogenous cells. These cells undergo mitosis and differentiate into microspore mother cells. The microspore mother cells undergo meiosis to form microspores still connected in tetrads. The tetrads separate into individual microspores which are released from the anther as mature pollen grains. Key tissues involved include the sporogenous tissue, tapetum, and anther wall layers.























































































