Downloaded 71 times
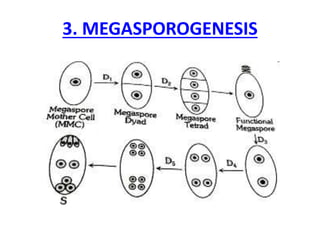
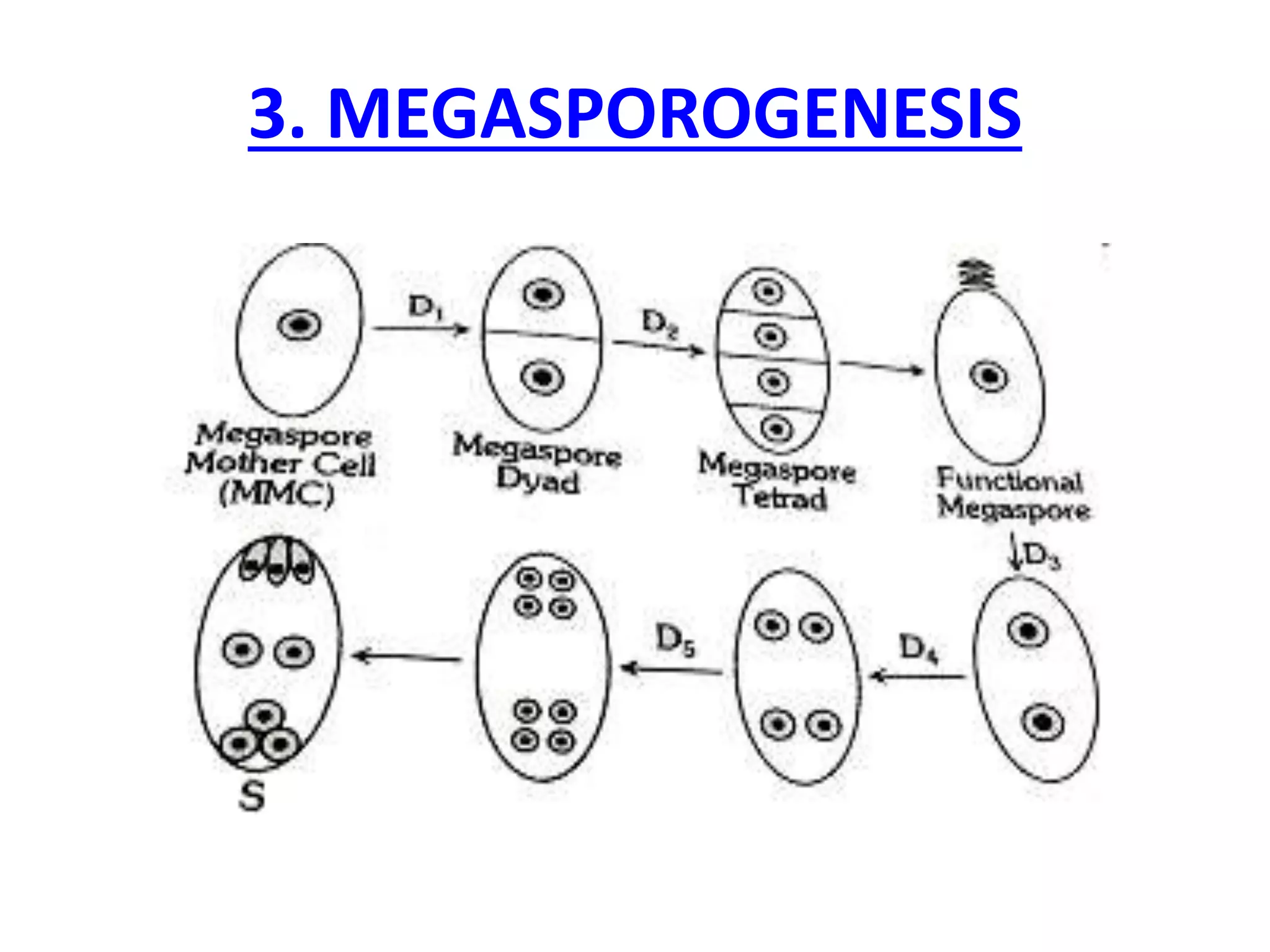




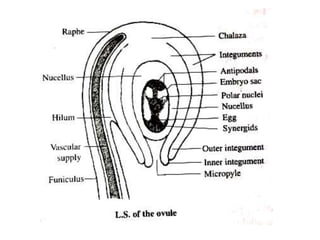
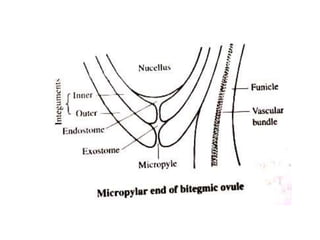







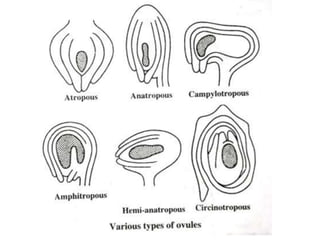







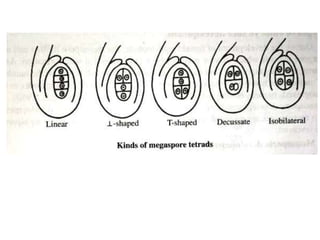
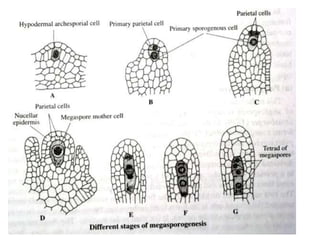











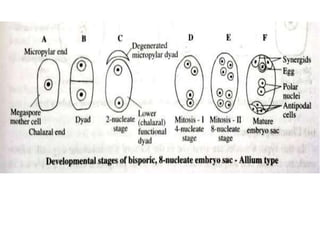



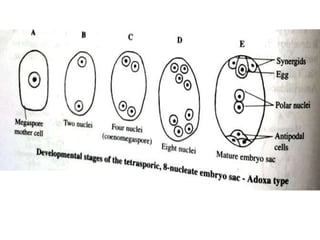







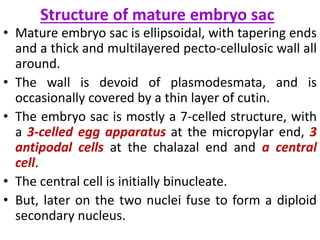

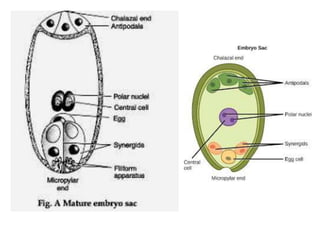


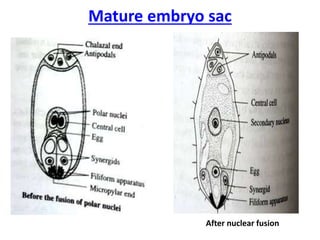




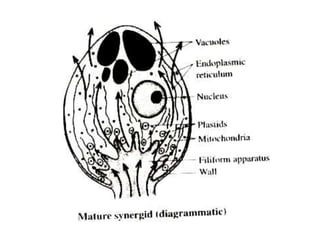





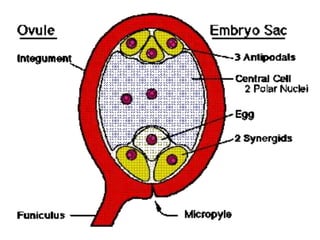



Megasporogenesis is the formation of megaspores in the megasporangium. This involves the development of the megaspore mother cell from the archesporial cell, which then undergoes meiosis to form four megaspores. One megaspore survives to form the embryo sac, which contains eight nuclei that develop into either a seven-celled structure in monosporic development types or other structures depending on the type of development. Ovules vary in structure depending on factors like the number of integuments and position of the micropyle.























































































