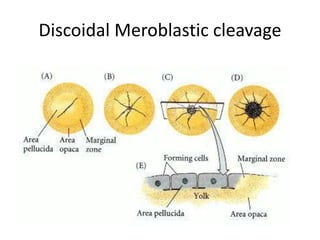
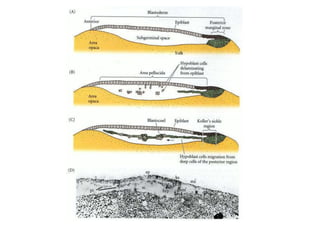
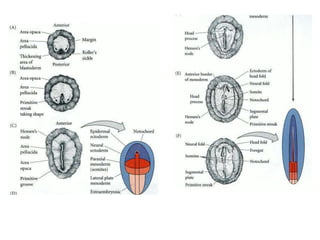
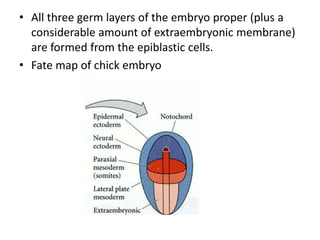
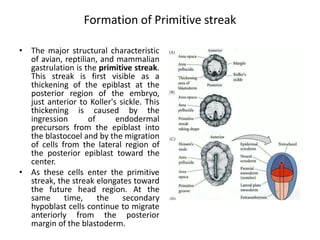
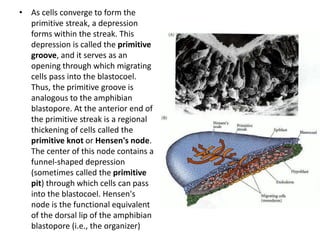
The document summarizes the development of a chick from an egg. It begins by describing the structure of the freshly laid hen's egg, including the large yolk, oval shape, membranes, and shell. It then discusses fertilization and cleavage, including discoidal meroblastic cleavage where cleavage is limited to the blastodisc atop the yolk. Next, it covers gastrulation processes like formation of the primitive streak and groove, migration of cells through the streak to form mesoderm and endoderm, and regression of the primitive streak.













































































![Polymer [ बहुलक ] Chemistry Notes PDF - Irfanullah Mehar - JJ Sir Chemistry.pdf](https://cdn.slidesharecdn.com/ss_thumbnails/polymerchemistrynotespdf-irfanullahmehar-jjsirchemistry-260210172118-3f9b37f7-thumbnail.jpg?width=640&height=640&fit=bounds)


















