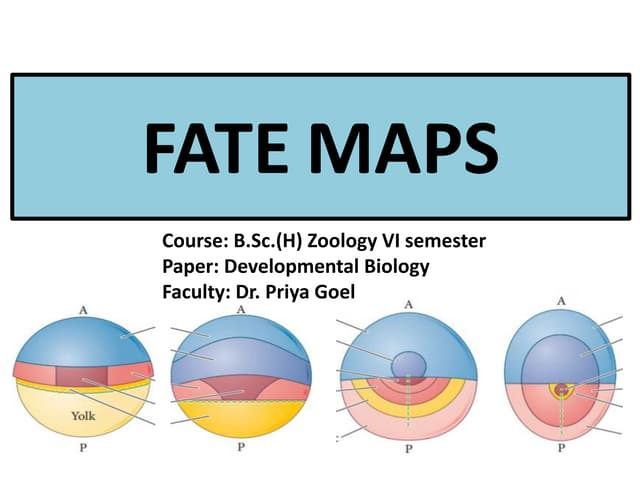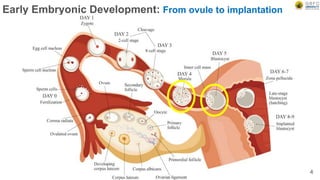
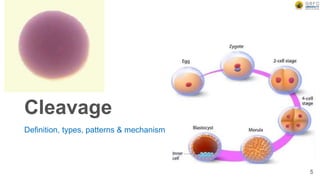
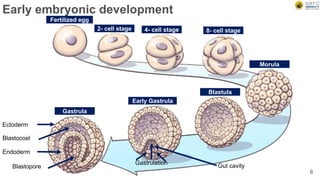











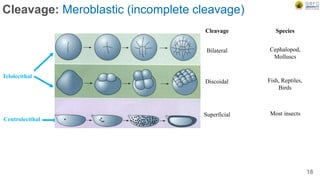
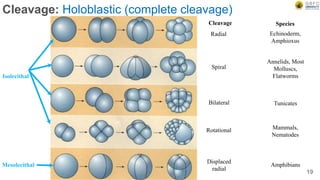





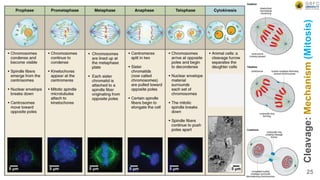





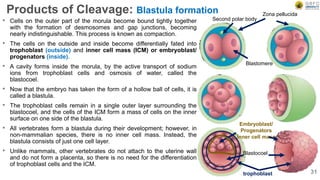
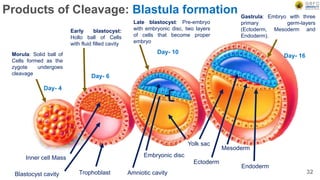



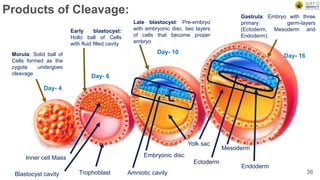
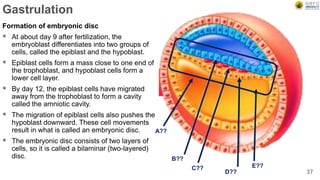

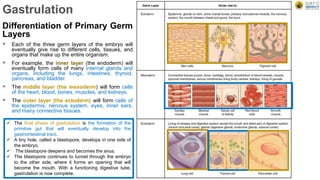



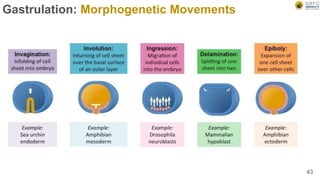
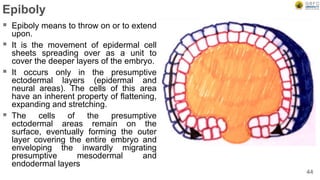
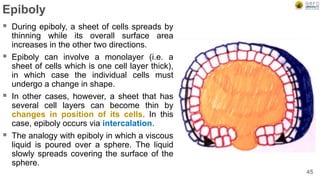

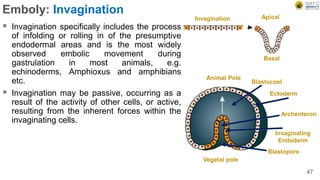
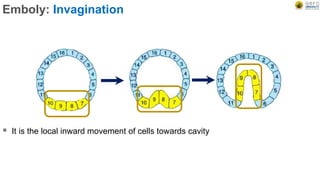
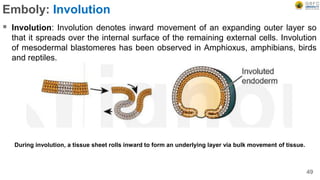
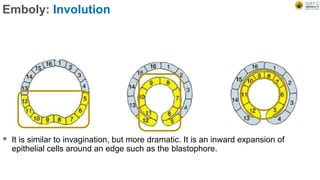
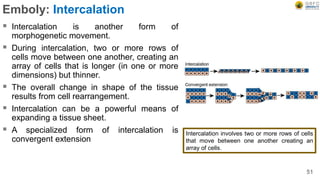
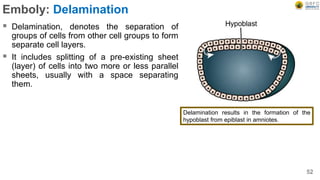
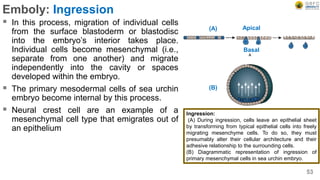












Early embryonic development involves a series of key stages: 1. Cleavage - The fertilized egg undergoes rapid, synchronized cell divisions without growth to form a solid ball of cells called a morula. 2. Blastulation - Cell divisions continue and a fluid-filled cavity called a blastocoel forms, establishing polarity and transforming the morula into a hollow ball of cells called a blastula. 3. Gastrulation - Cells migrate and rearrange through morphogenetic movements to form the three primary germ layers - ectoderm, endoderm, and mesoderm - establishing the body plan of the embryo.