

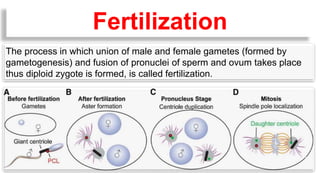
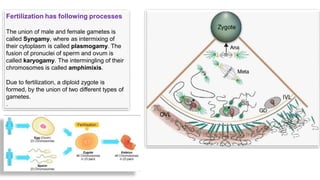

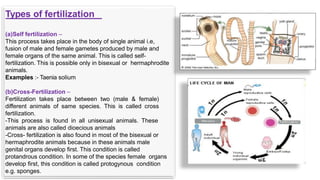

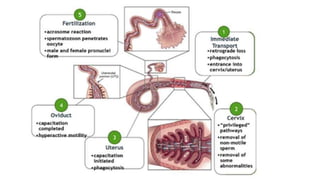




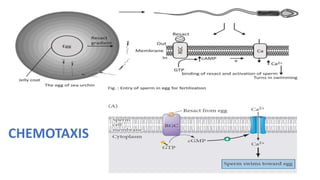

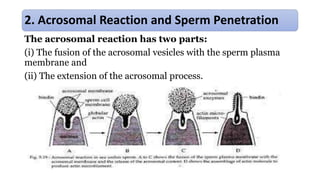


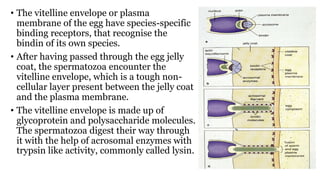

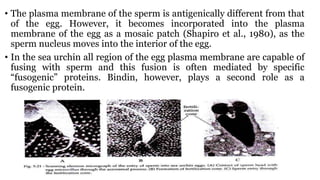






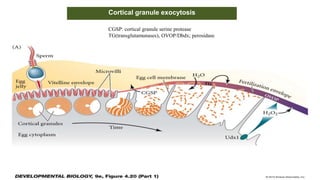
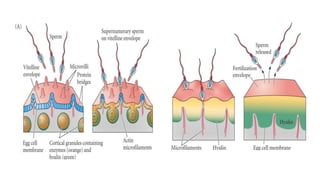
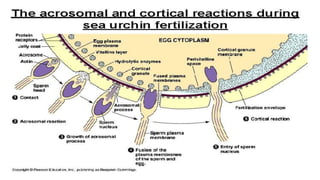
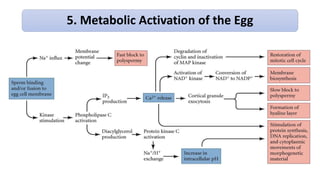
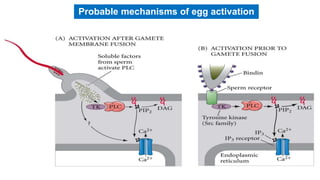

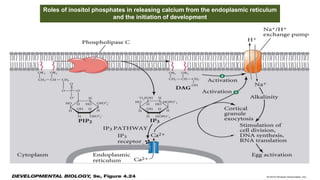


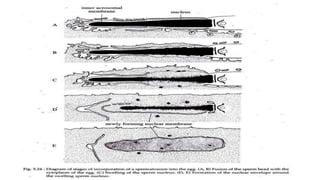
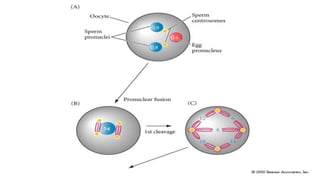


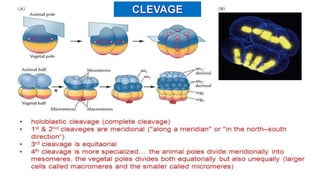
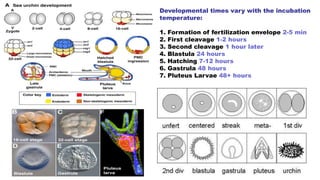



Fertilization in sea urchins involves several key steps: 1) Sperm are attracted to eggs via chemotaxis using peptides like resact. 2) The acrosomal reaction allows sperm to penetrate the egg jelly and bindin aids binding to the egg. 3) Prevention of polyspermy involves a fast block changing membrane potential and slow block from cortical granule exocytosis. 4) Metabolic activation and pronuclear fusion within the egg forms a zygote, completing fertilization.











































































