Downloaded 320 times













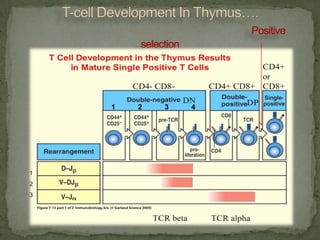

























The document discusses the mechanisms of T-cell and B-cell development and tolerance in the immune system, focusing on the processes that enable high specificity and sensitivity without self-attack. It details T-cell maturation in the thymus, including positive and negative selection, the roles of different T-cell lineages, and the influence of the autoimmune regulator (AIRE) protein. Additionally, it covers B-cell development, the importance of immunoglobulin expression, and how receptor editing can prevent self-reactivity.











































![Blood group.007[1]](https://cdn.slidesharecdn.com/ss_thumbnails/bloodgroup-140326055444-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)











































