Downloaded 78 times










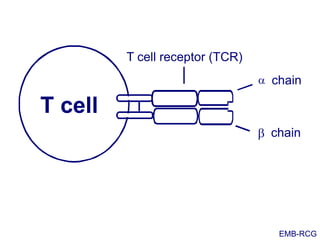



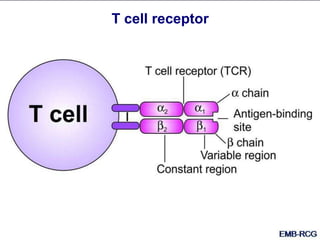






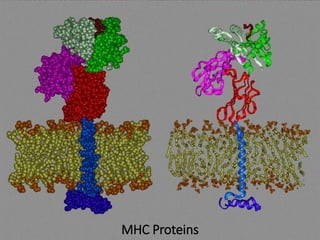





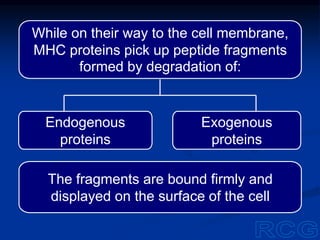


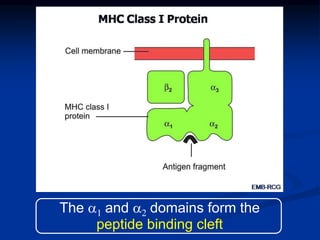
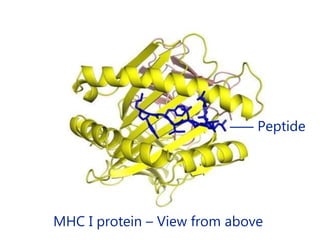


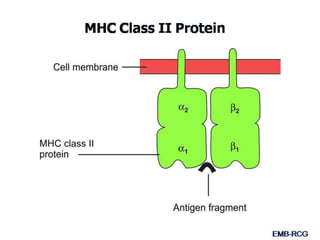
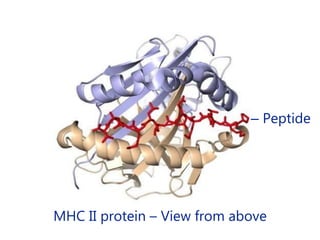







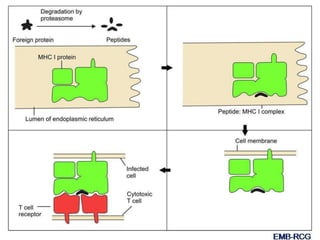





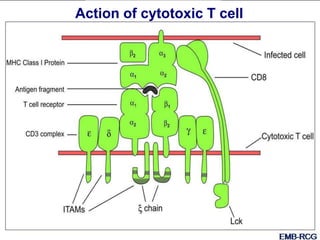







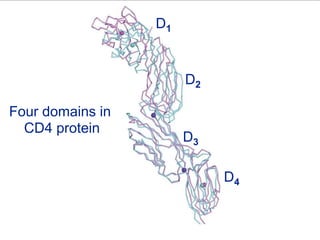

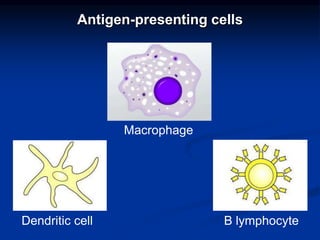



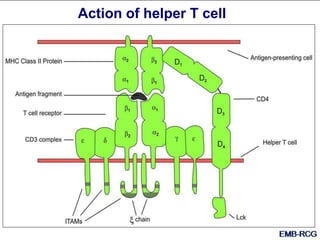

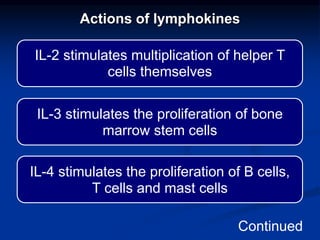





















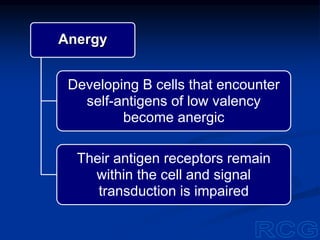






















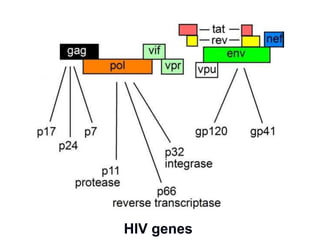




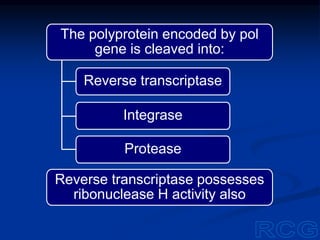



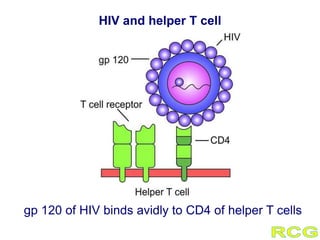





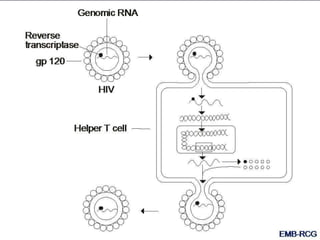

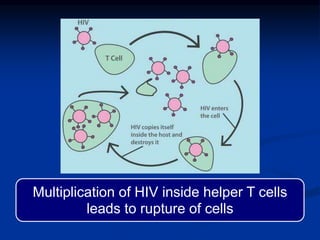




This document summarizes cell-mediated immunity. It discusses how T lymphocytes are responsible for cell-mediated immunity, with cytotoxic T cells destroying infected cells and helper T cells activating other immune cells through cytokine release. Antigens from intracellular pathogens are presented on MHC class I to cytotoxic T cells, while extracellular pathogens are presented on MHC class II to helper T cells. The document also describes T cell receptor structure and diversity, antigen recognition, and the mechanisms of cytotoxic T cell and helper T cell activation.























































































