


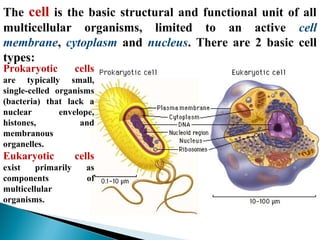
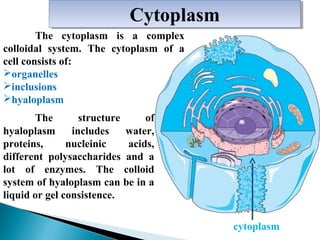
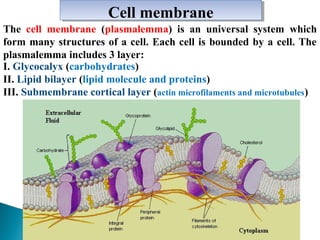
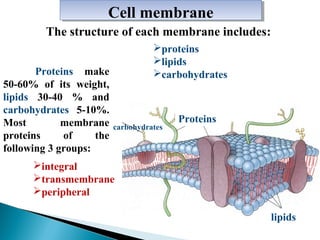
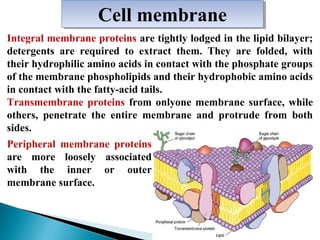
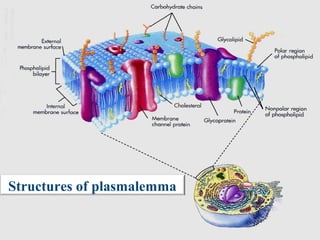
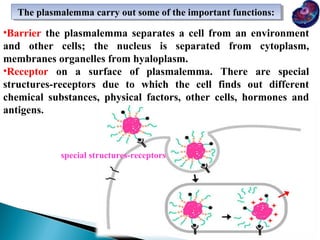
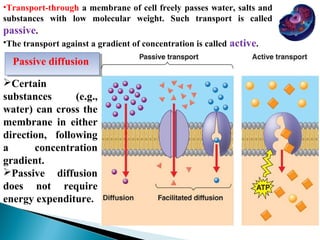
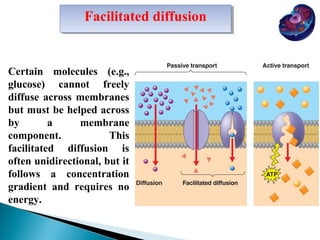
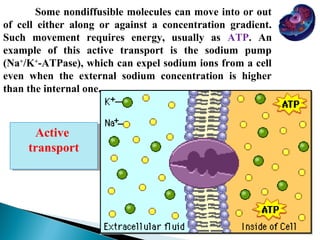
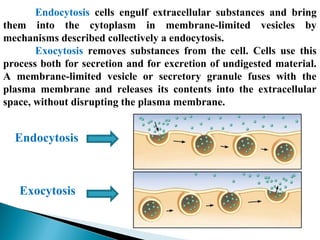
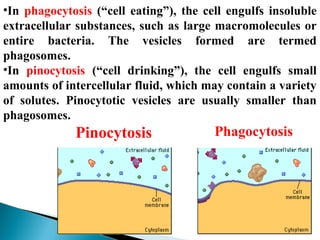
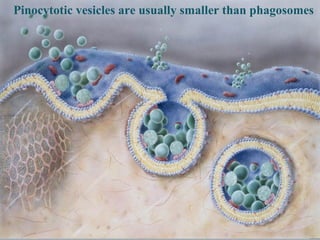
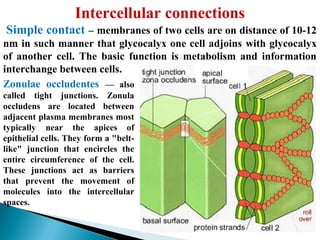
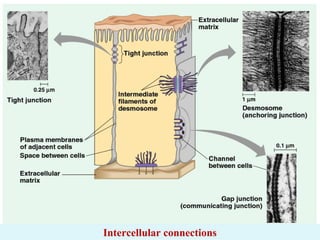
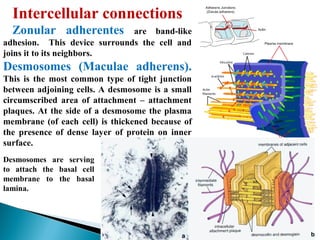
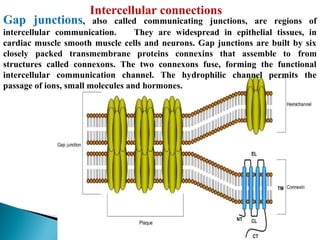
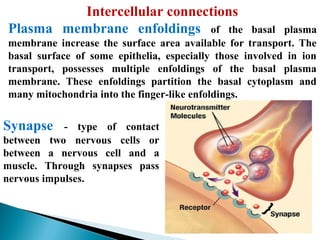


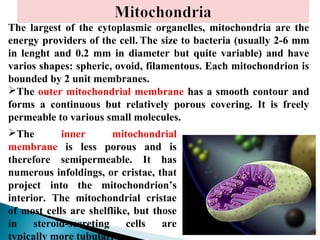
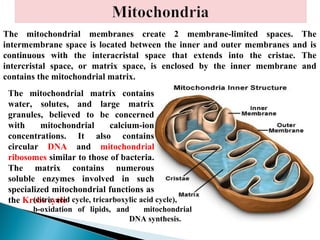
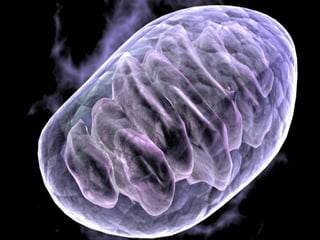

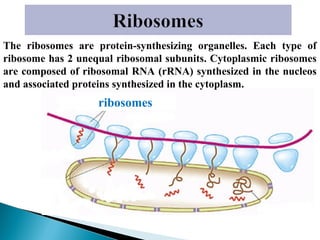
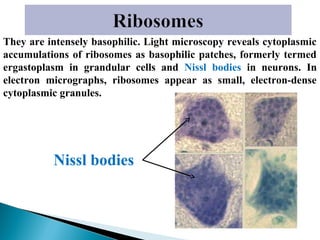
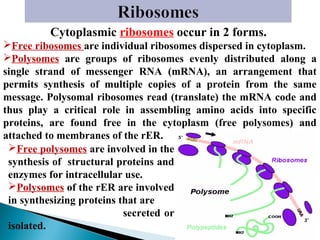
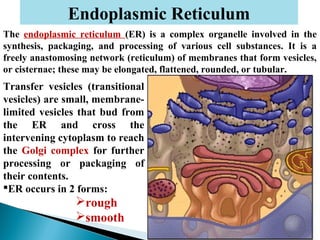
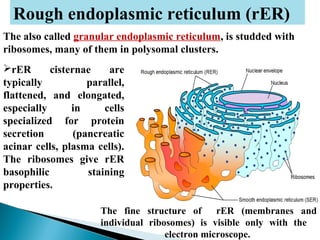

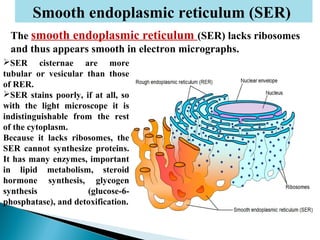
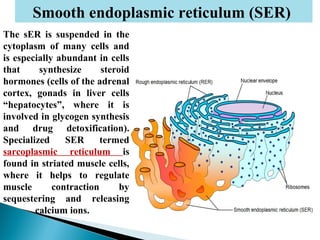
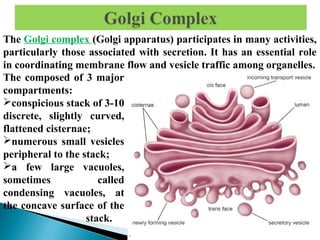
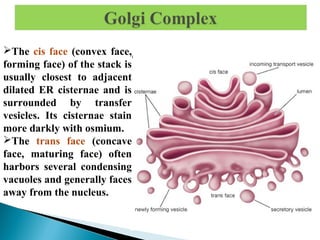



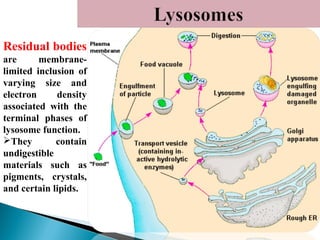
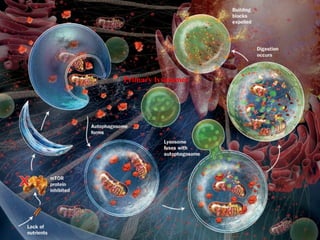

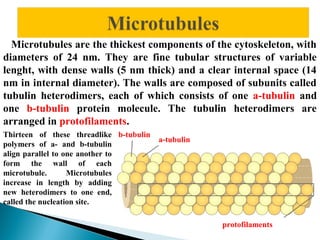

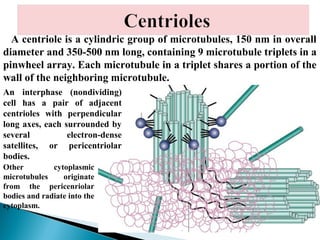
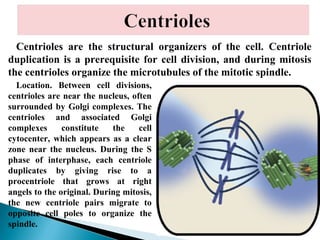

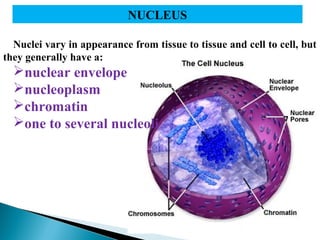
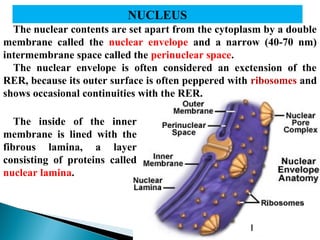
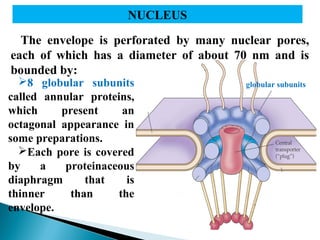
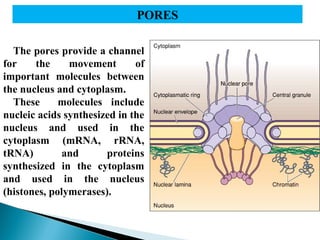
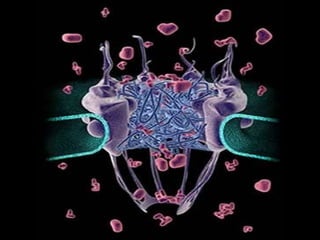
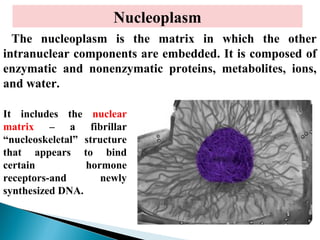
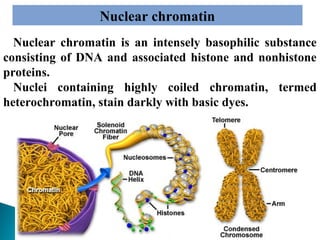

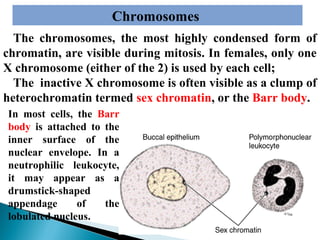
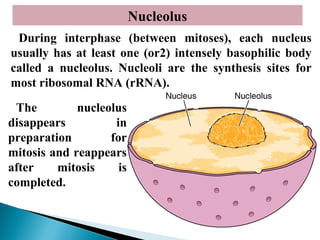
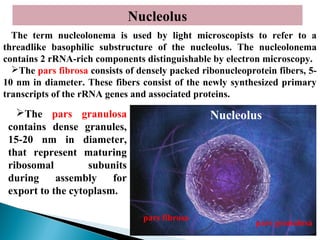
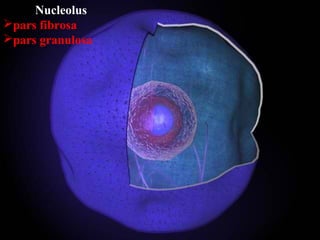


- Histology is the study of tissues and how they form organs. It examines tissues at a microscopic level. - Cytology is the study of cells. There are two main cell types - prokaryotic cells which are small and lack organelles, and eukaryotic cells which exist primarily in multicellular organisms and have organelles. - The cell membrane forms the boundary of the cell and regulates what enters and exits. It contains proteins, lipids, and carbohydrates. It performs important functions like acting as a barrier, having receptor sites, and facilitating transport processes into and out of the cell.




















































![FINAL CELLS ANATOMY (1) [Auip8uui;oy9'yiuy79y08ugtttosaved].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/finalcellsanatomy1autosaved-250115165251-ffe2d15c-thumbnail.jpg?width=640&height=640&fit=bounds)








































