Download as PDF, PPTX


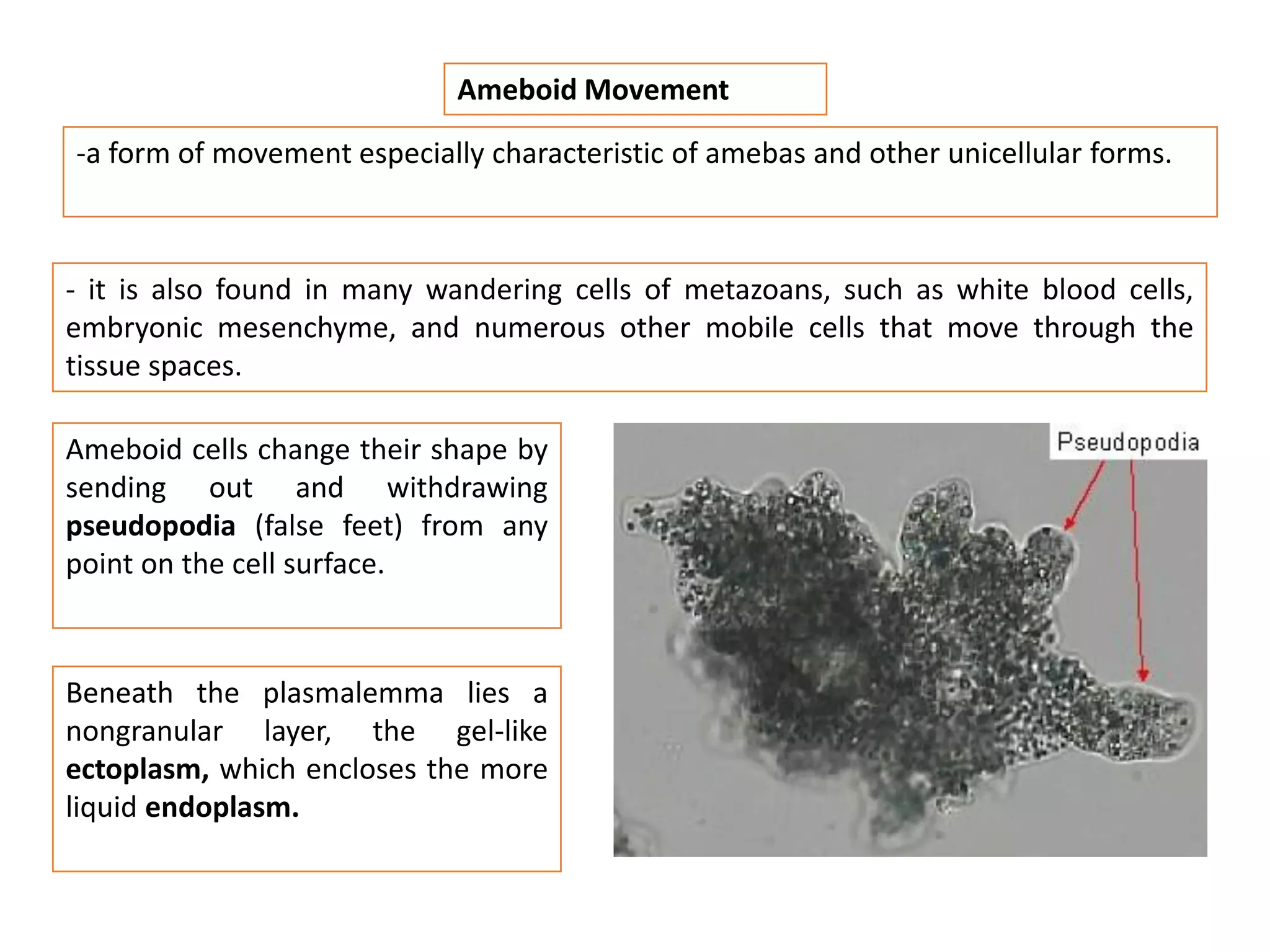
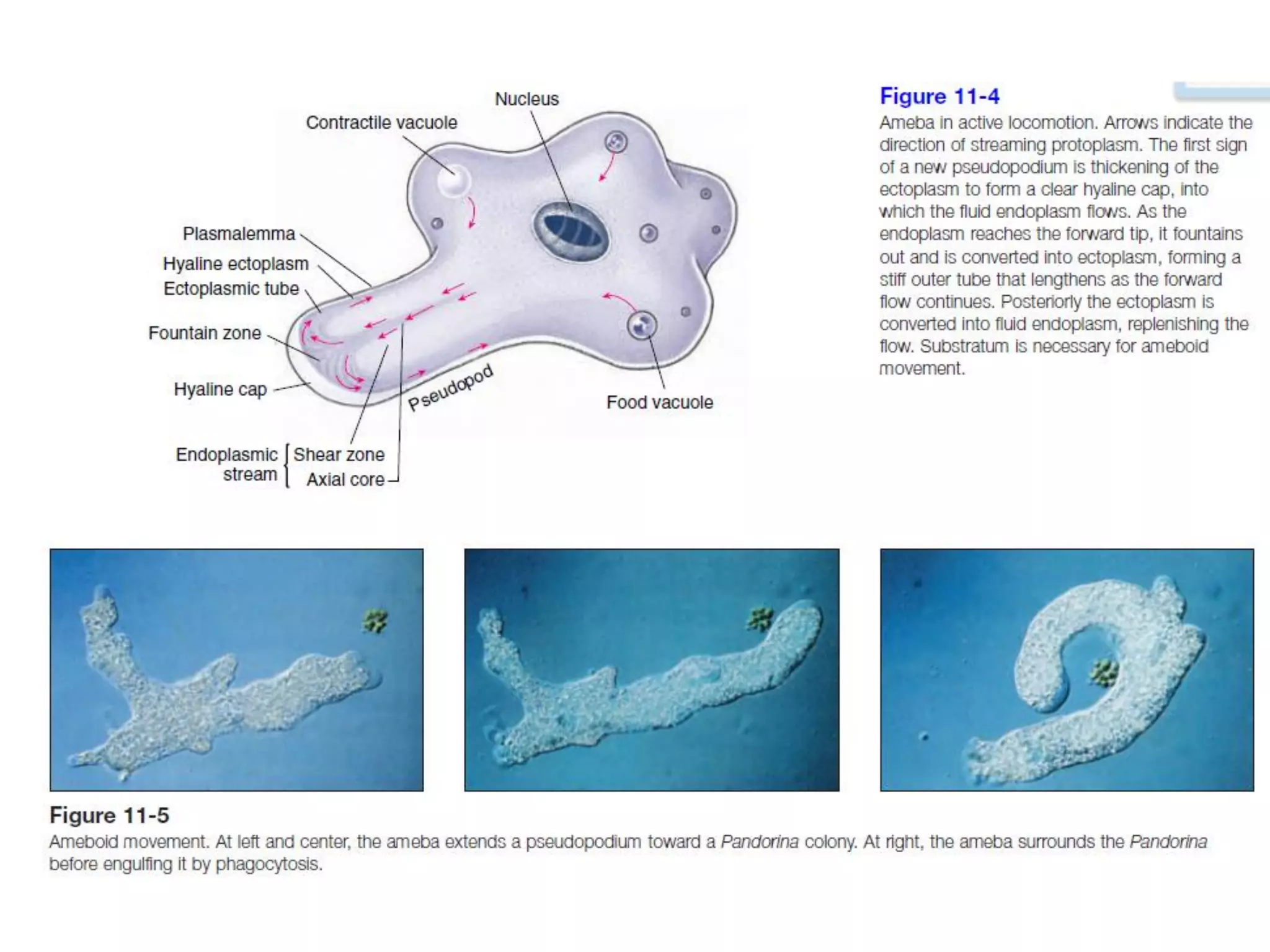
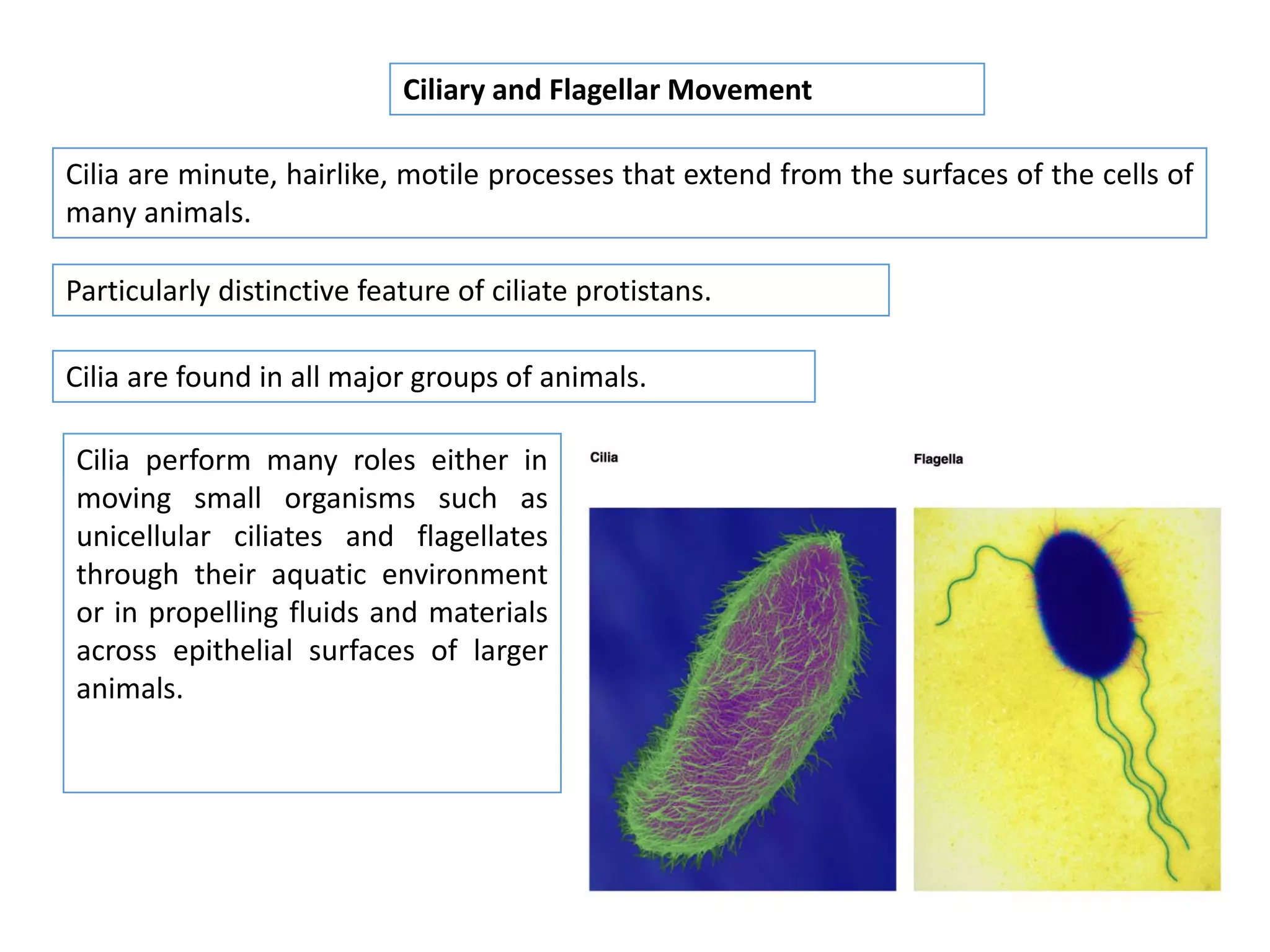
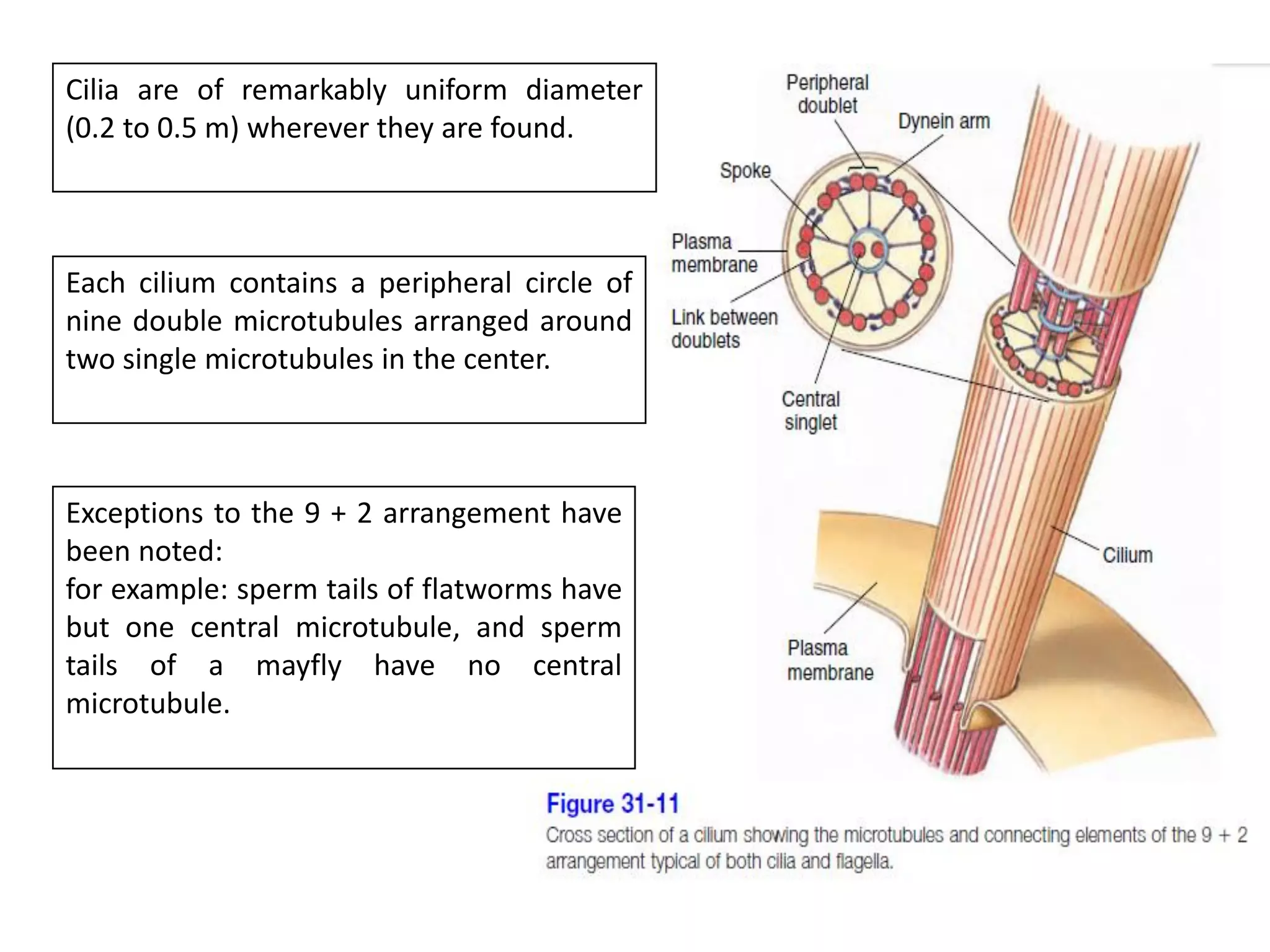
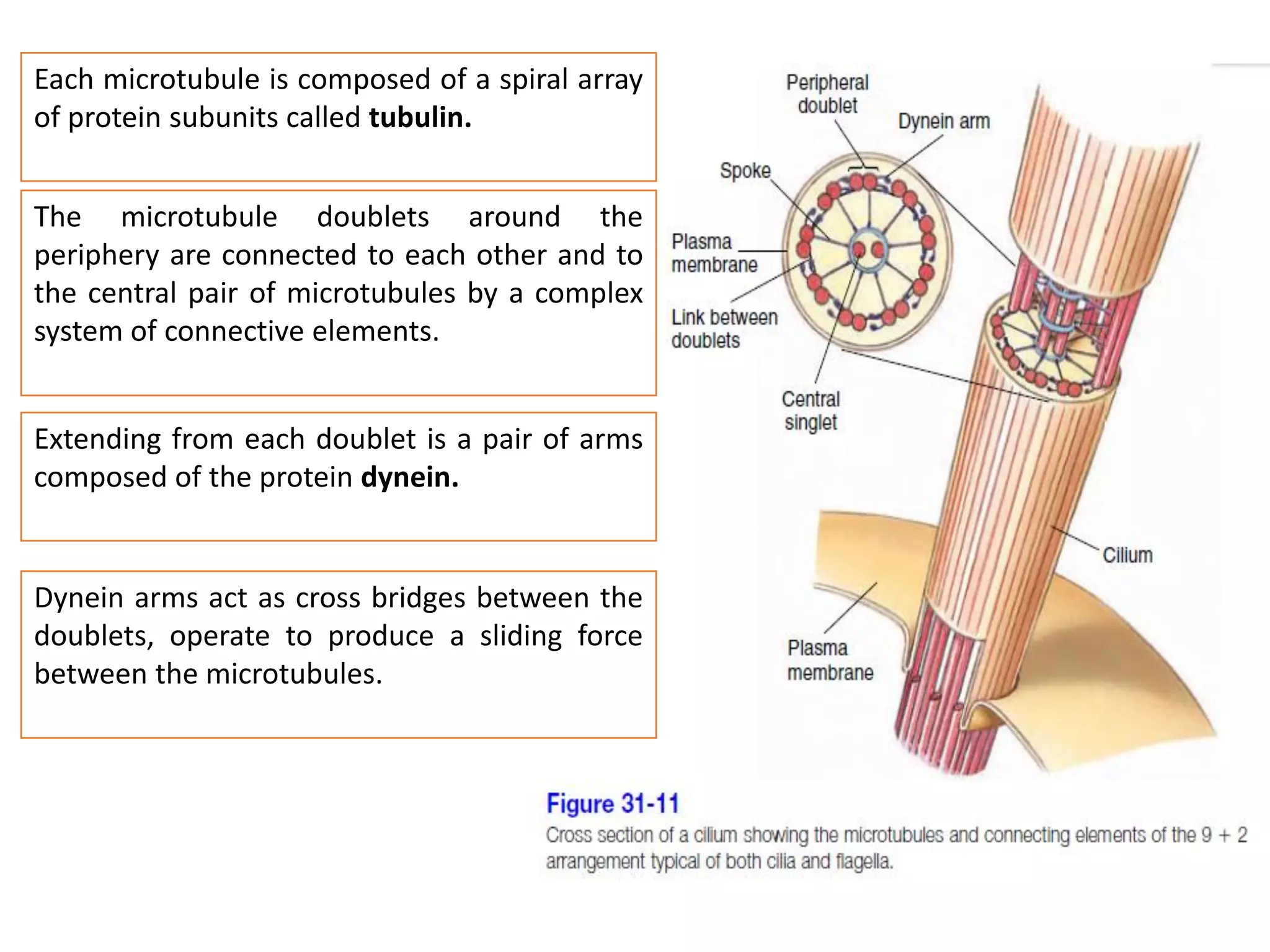
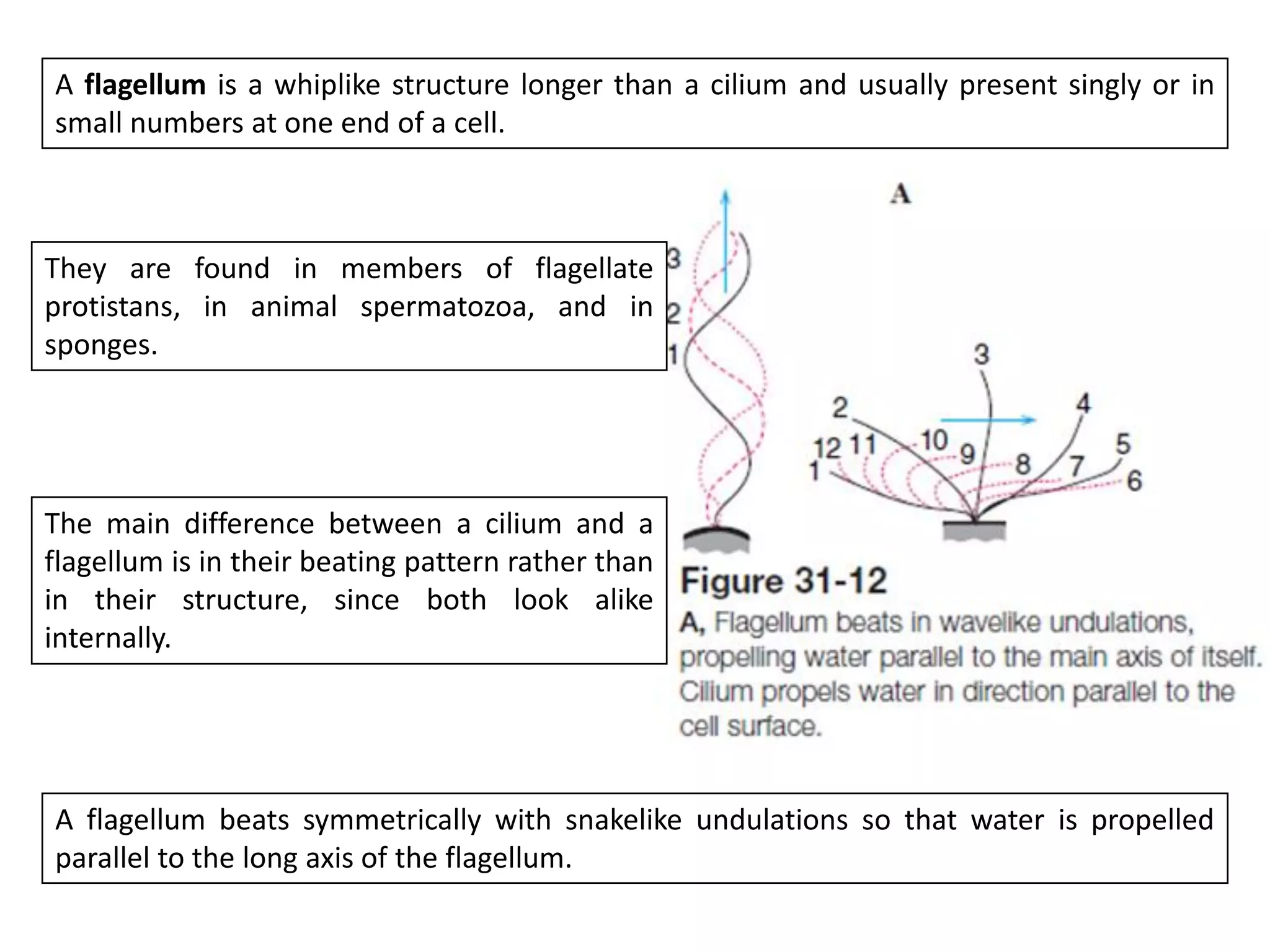
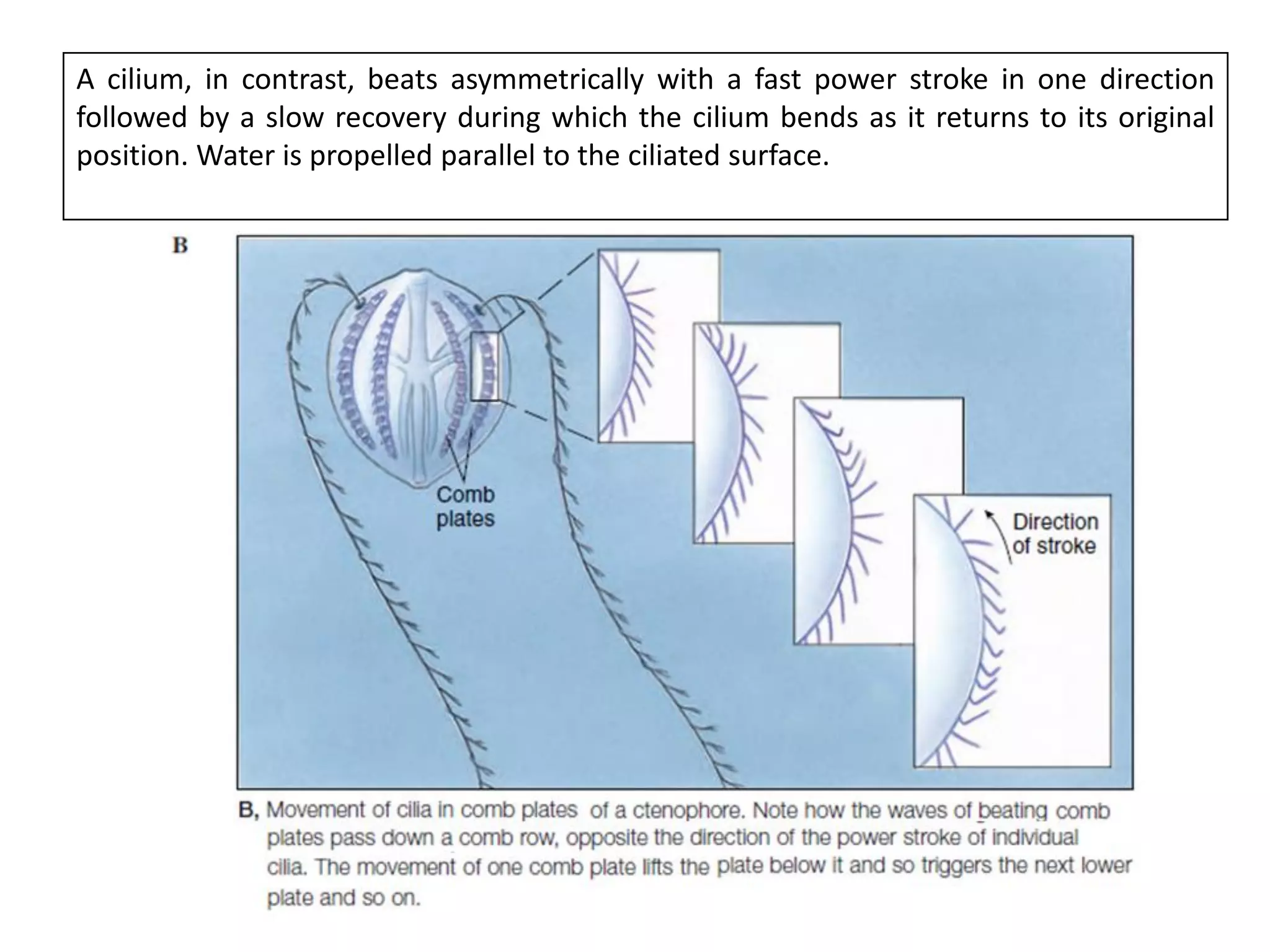
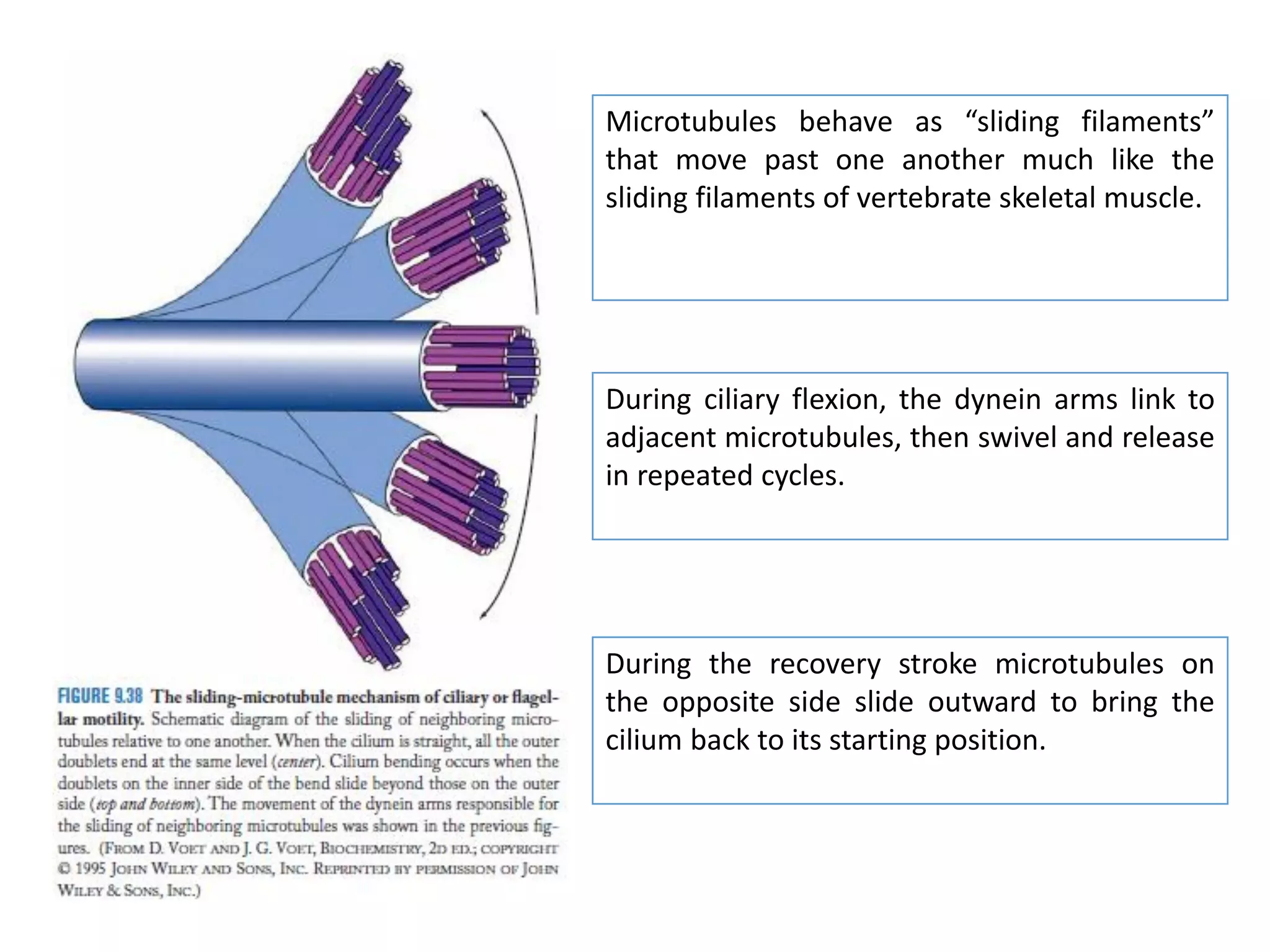
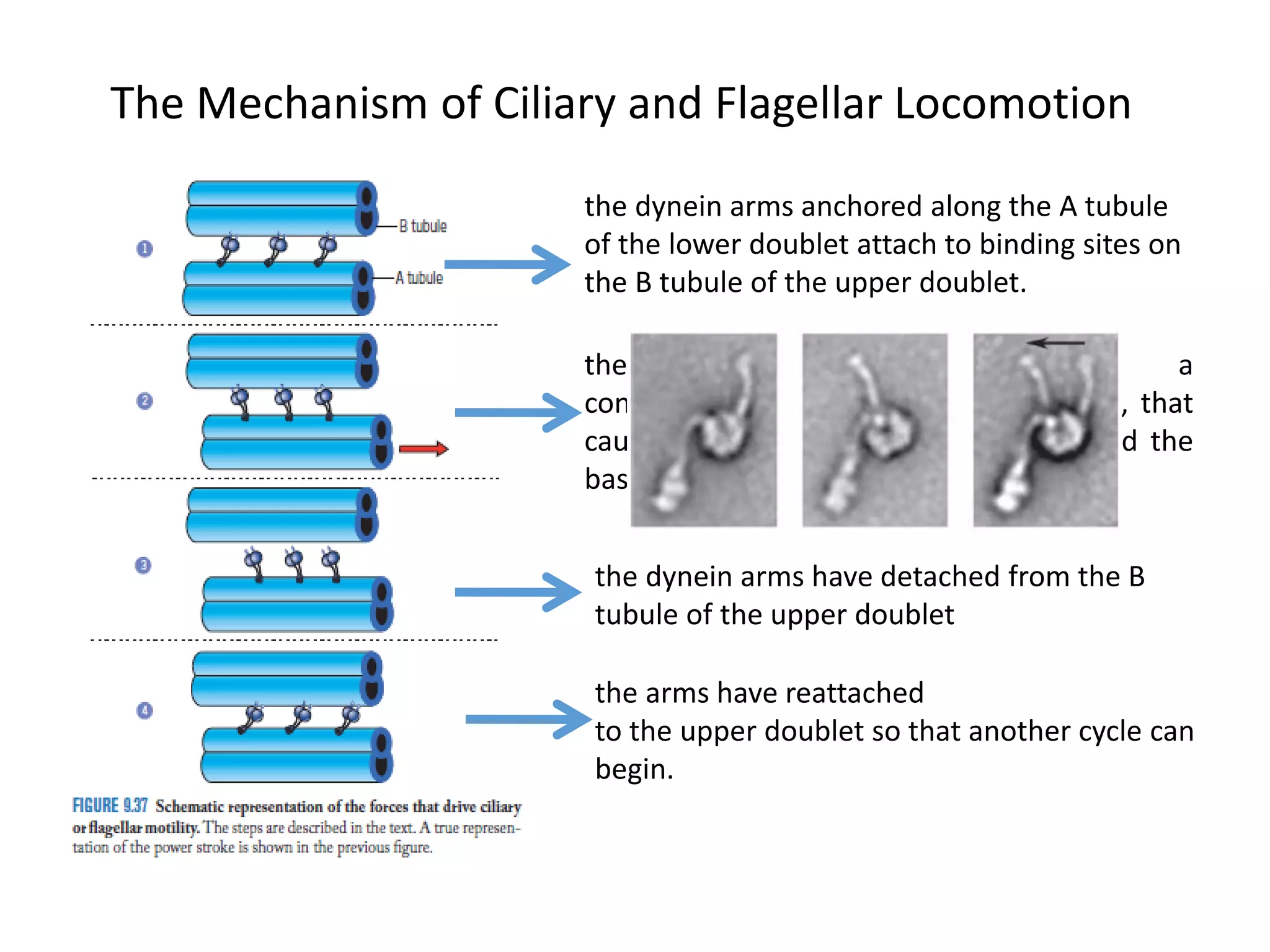


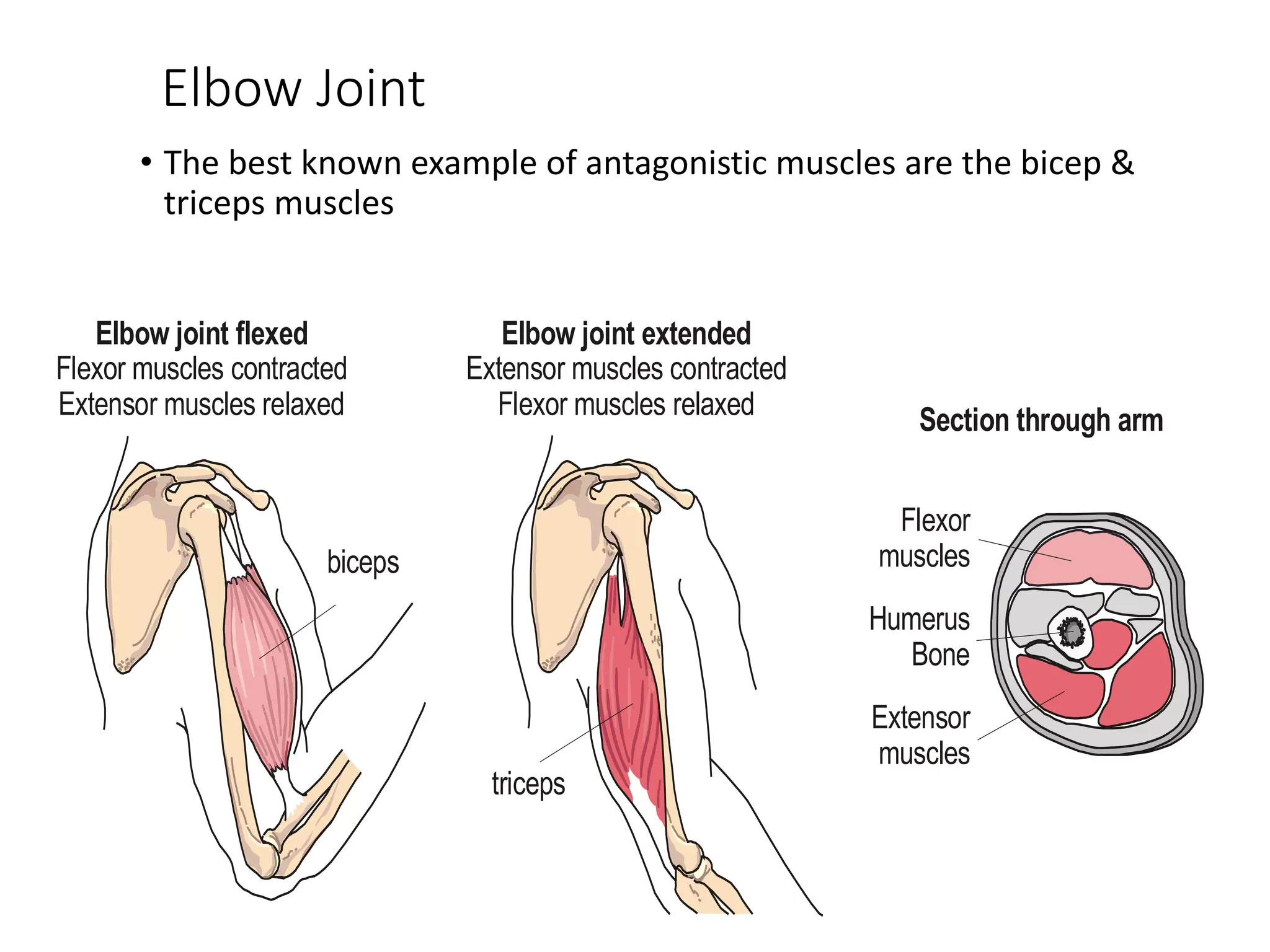
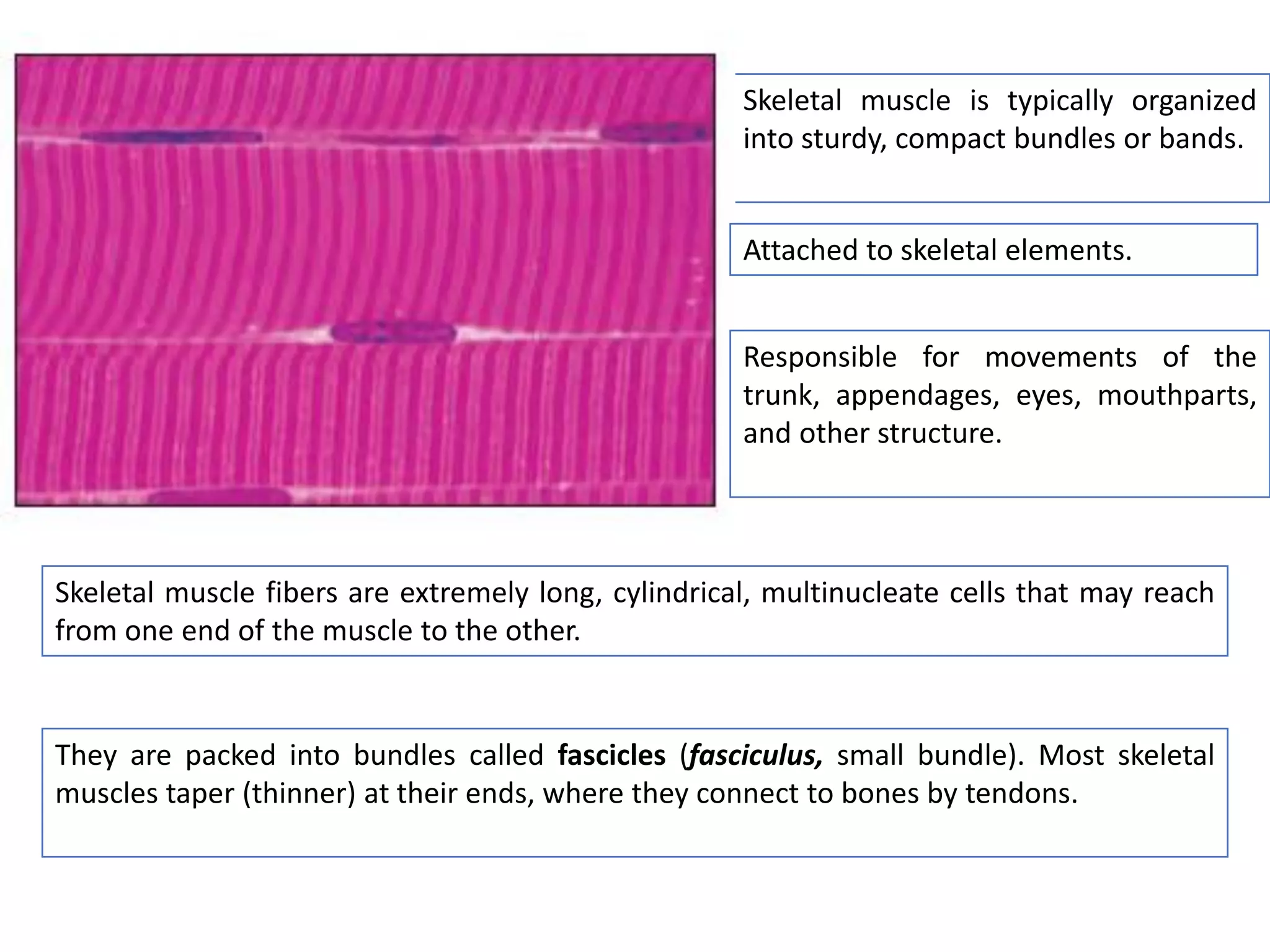

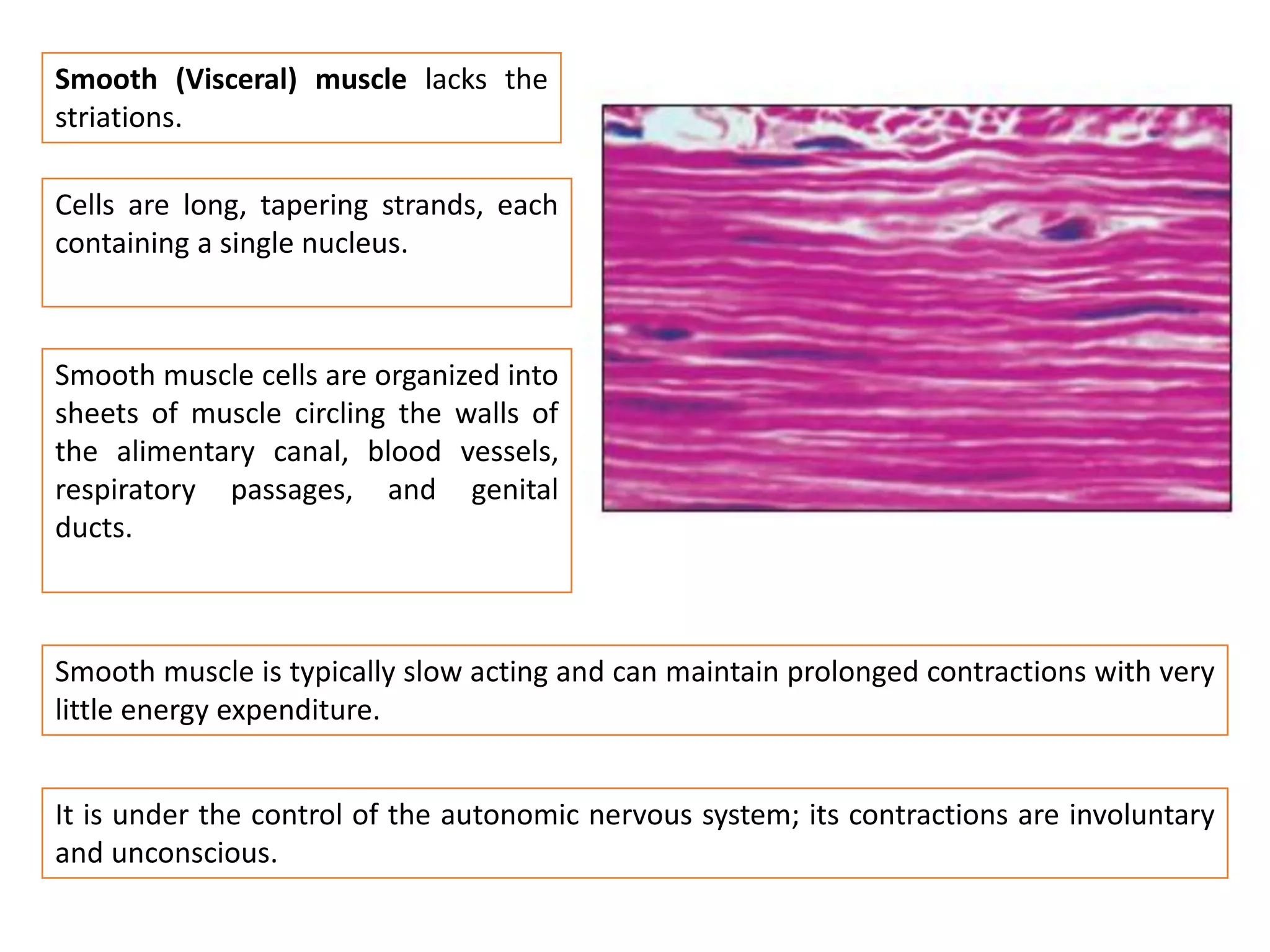
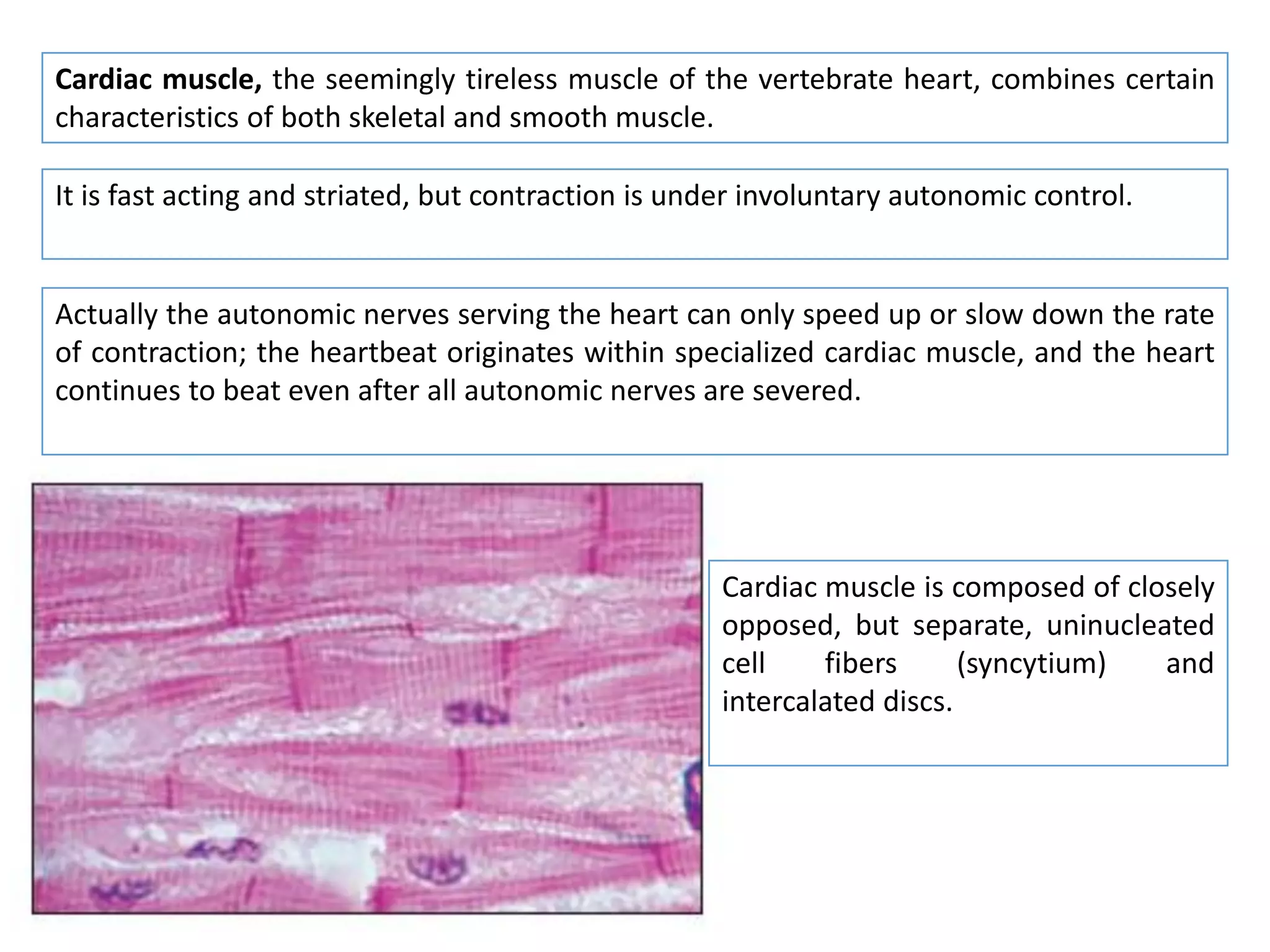

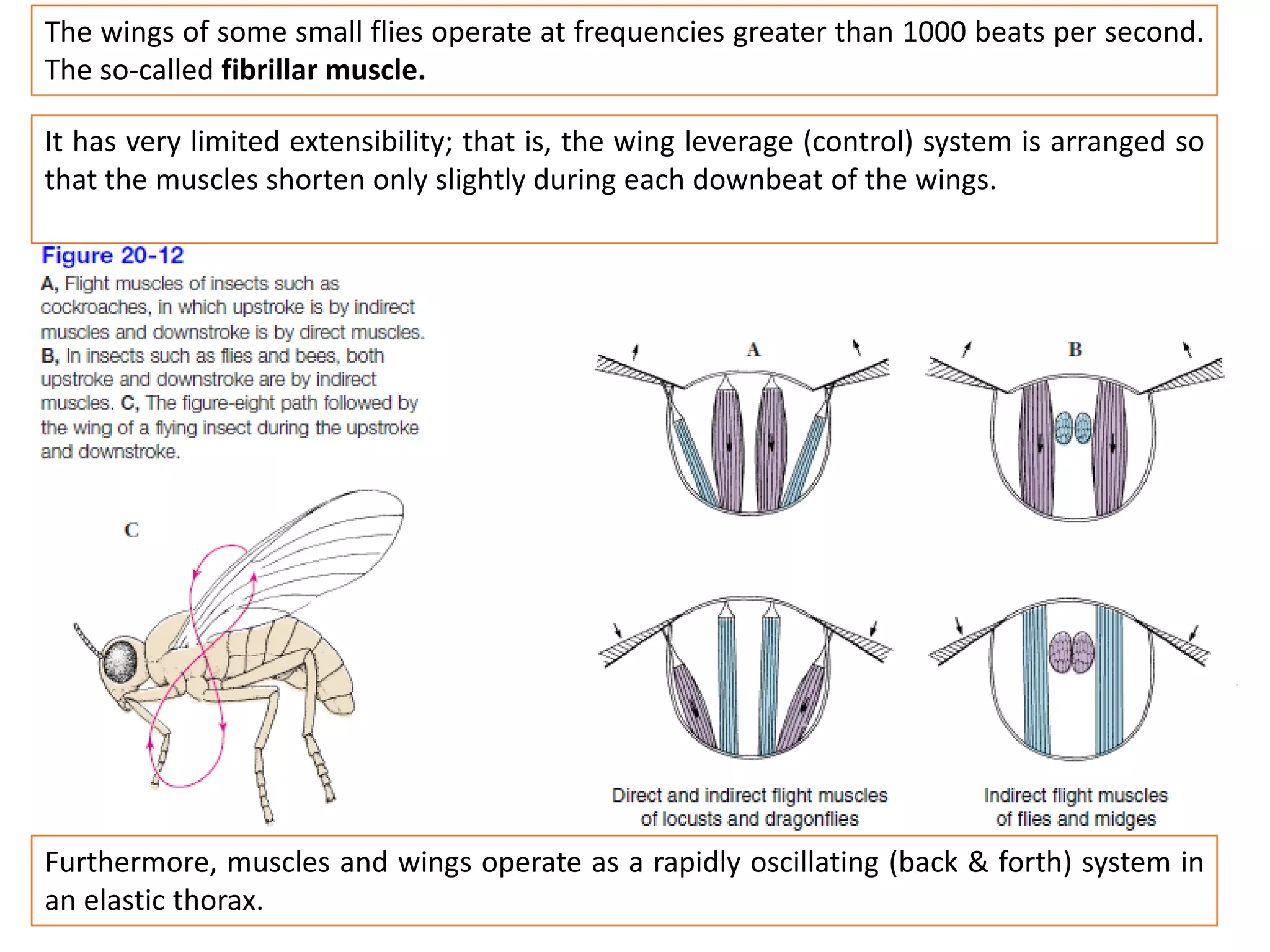

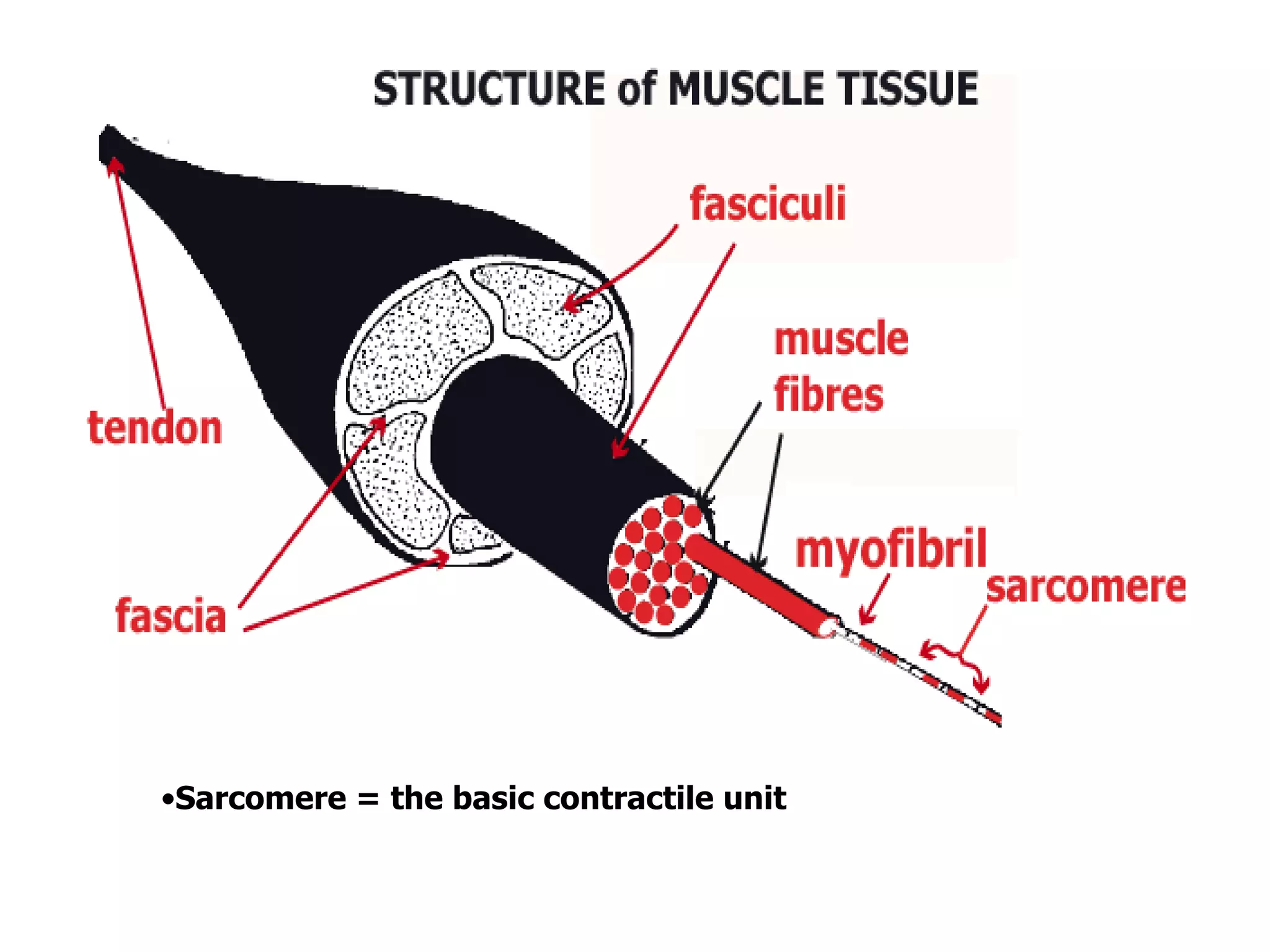
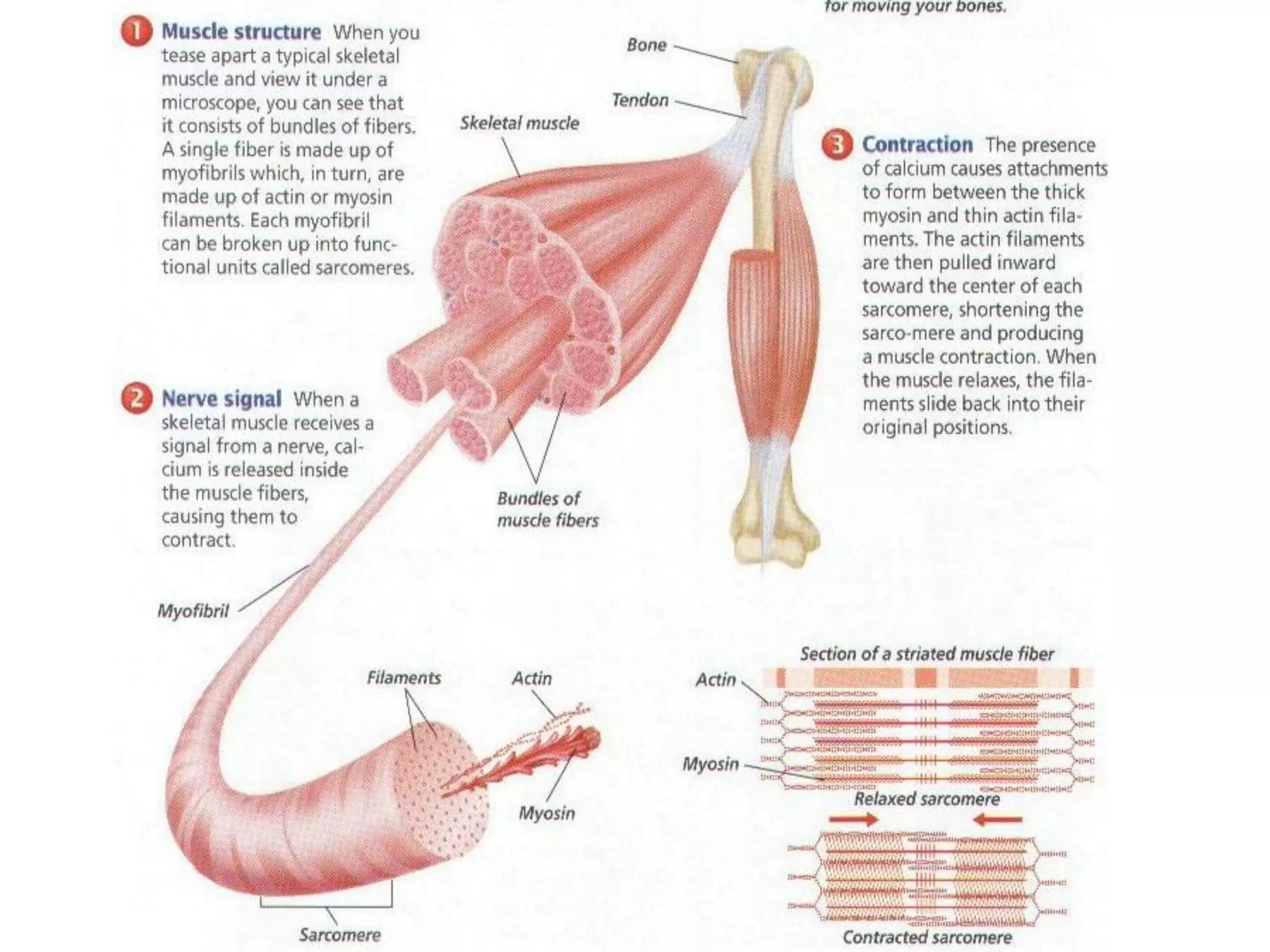
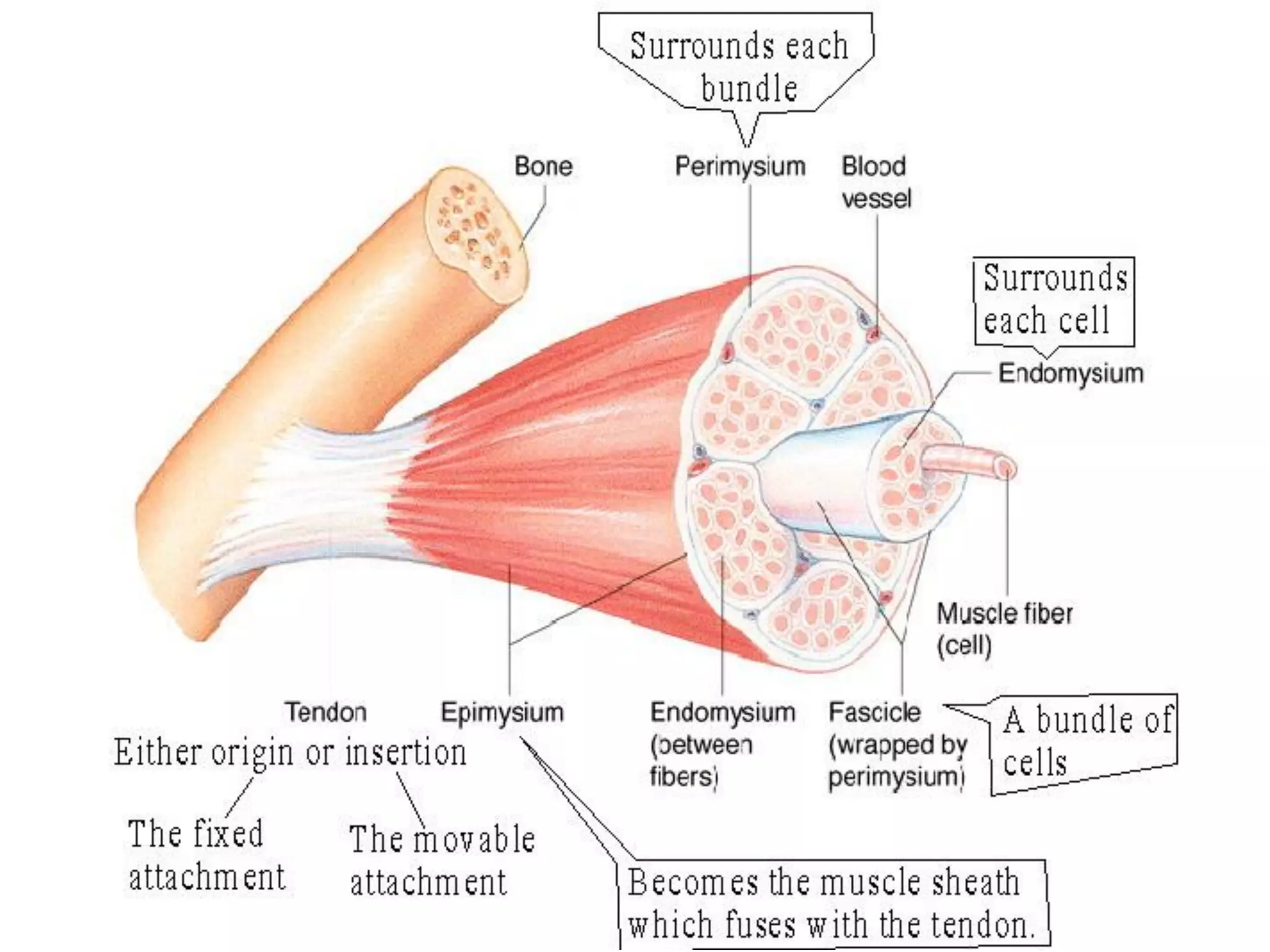
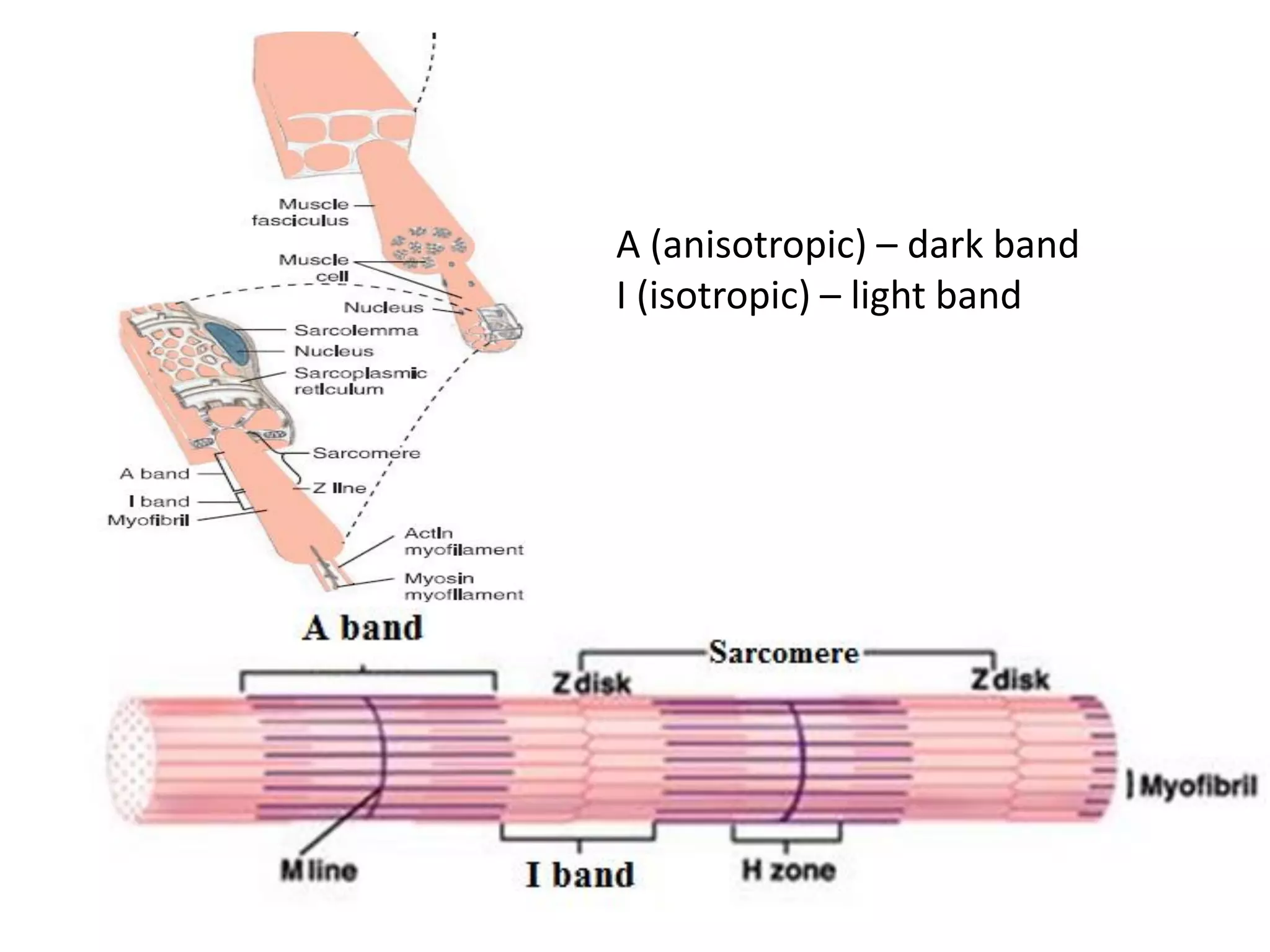
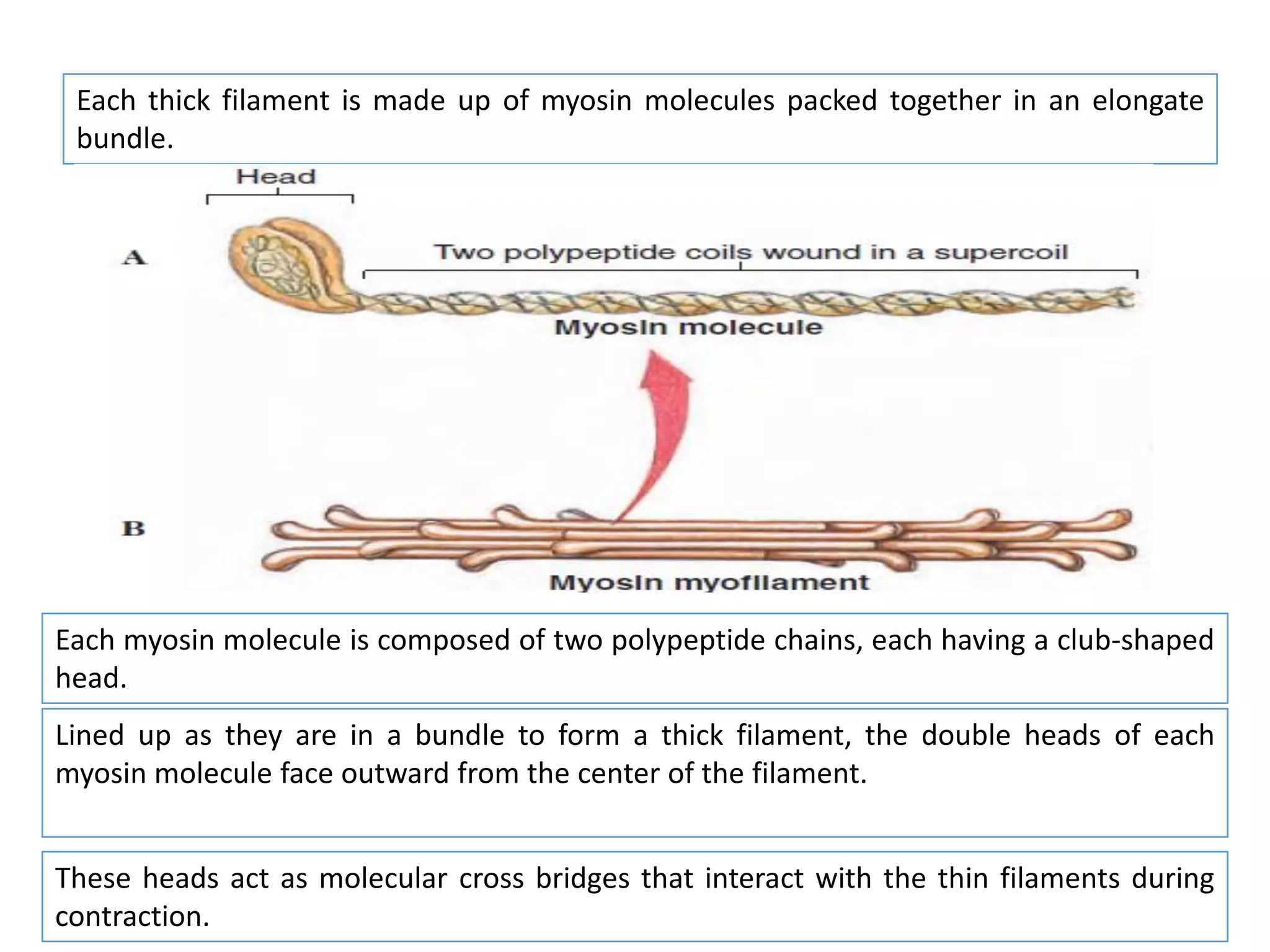
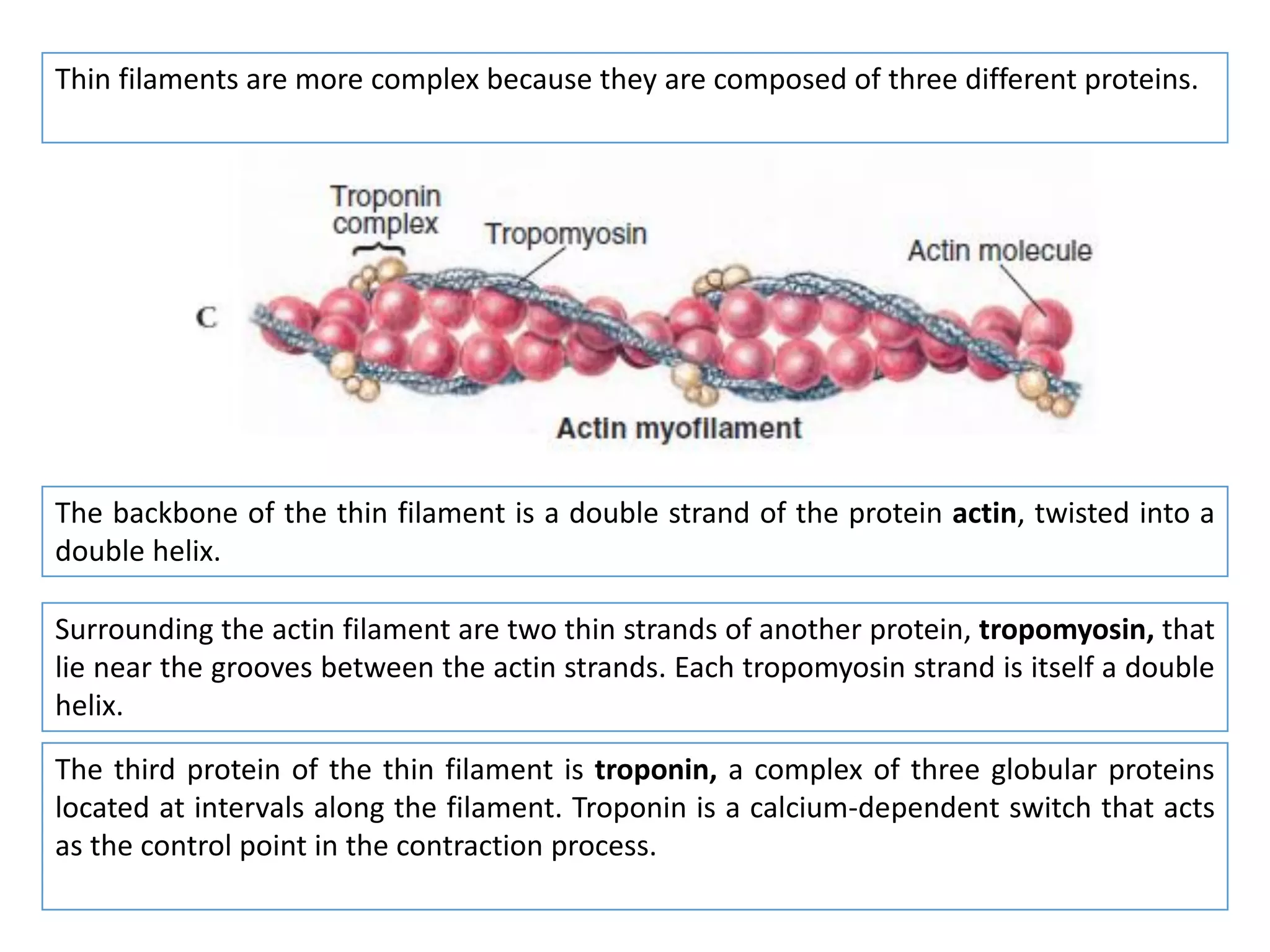
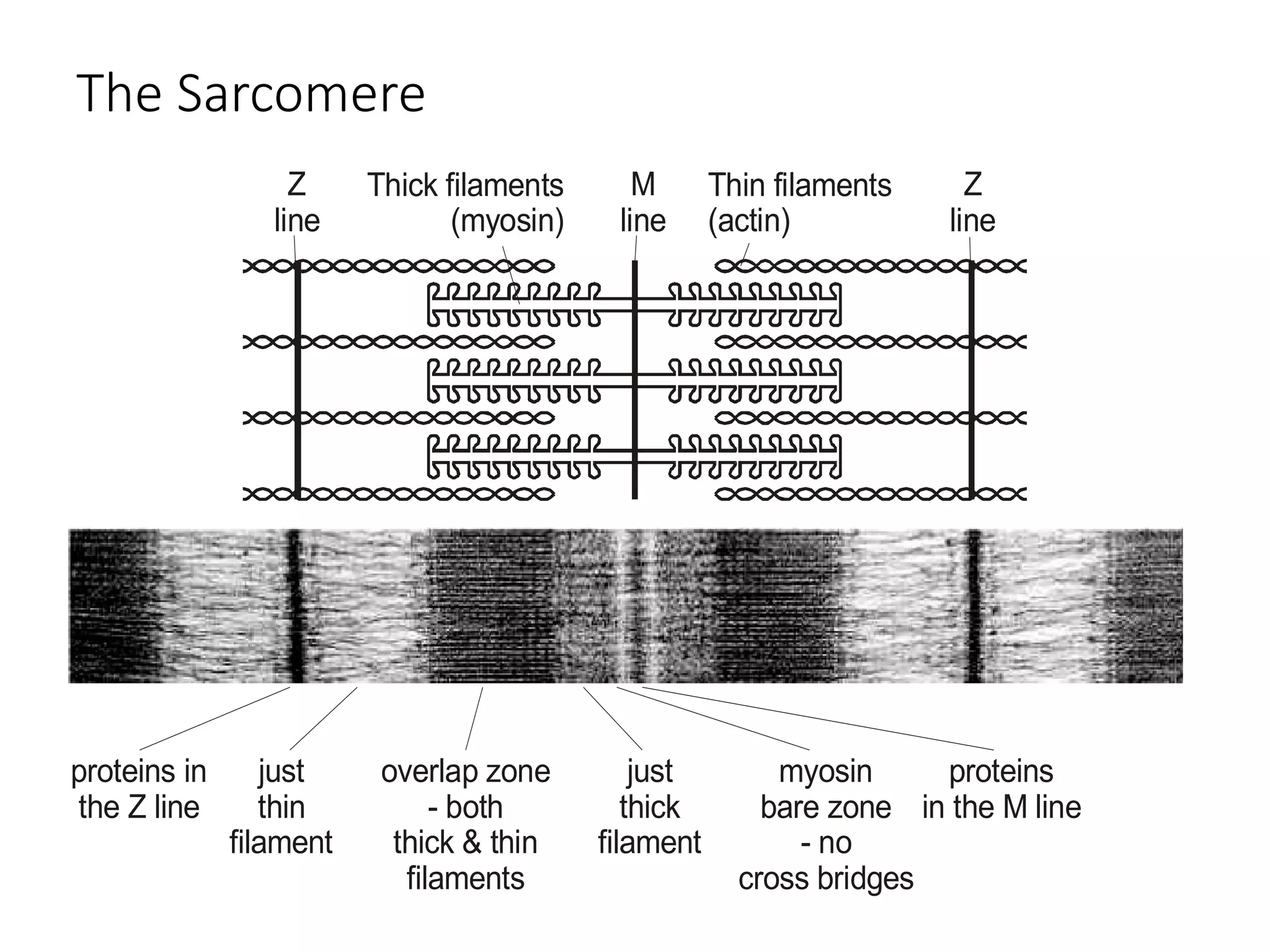
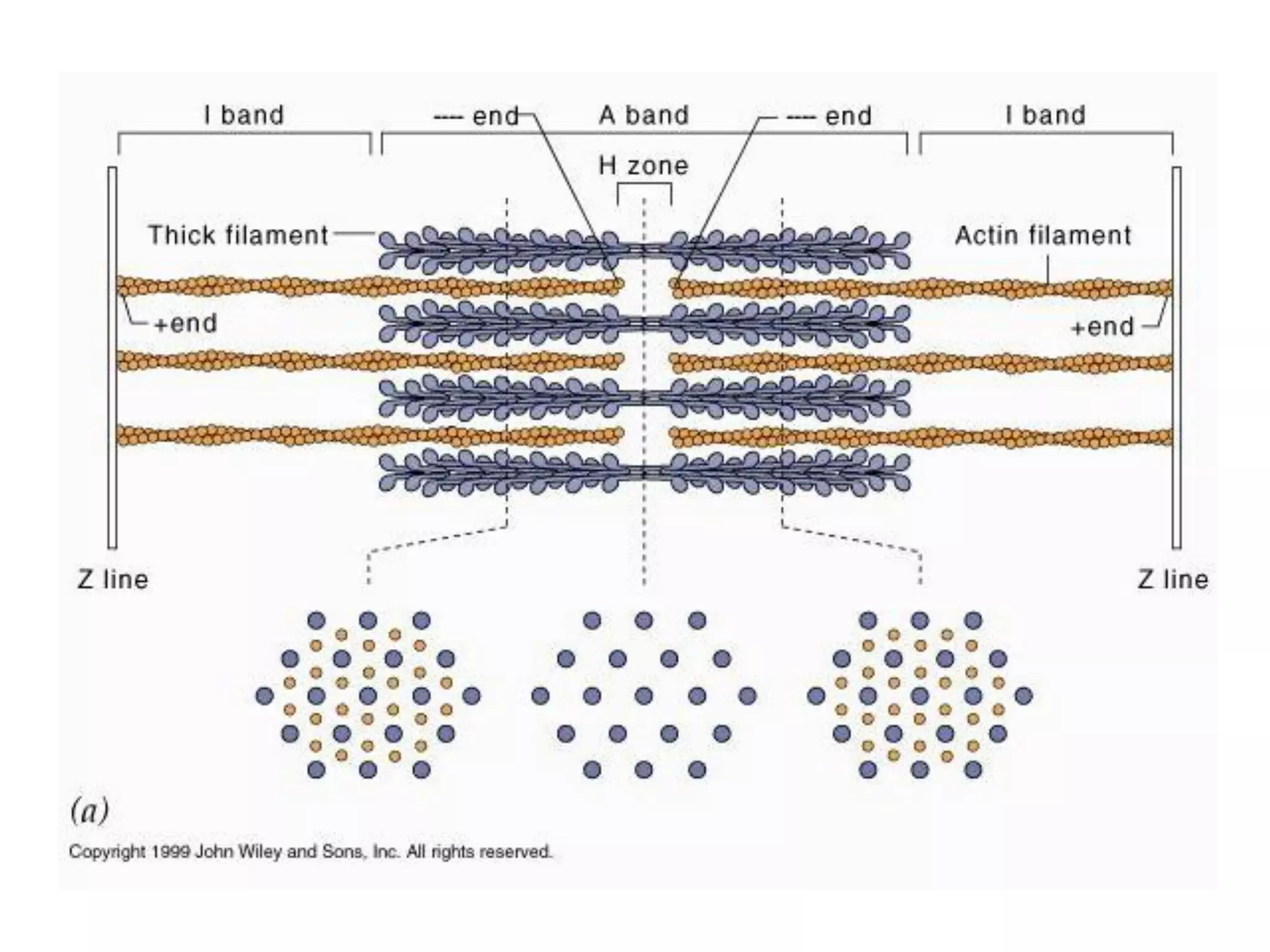
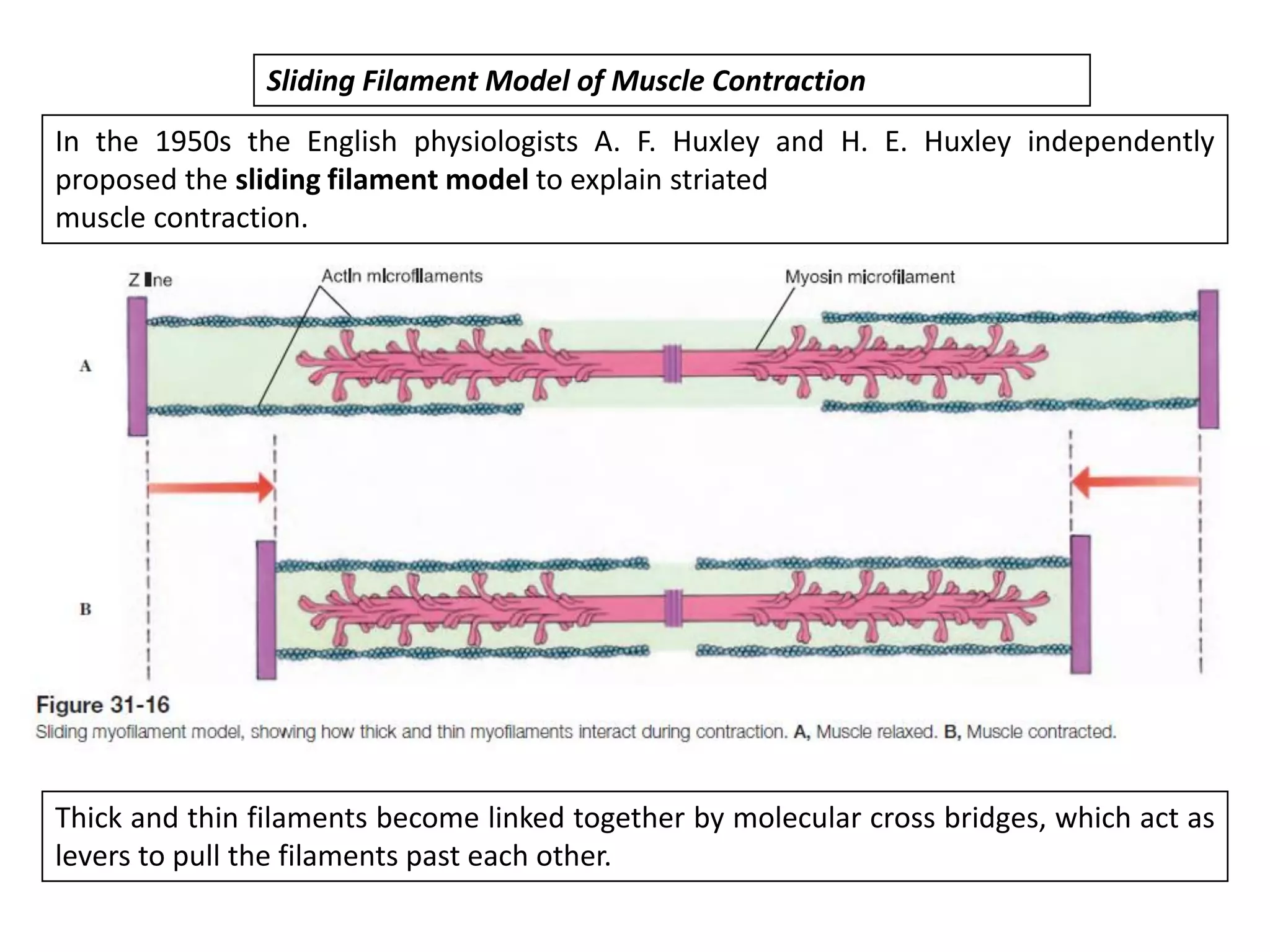
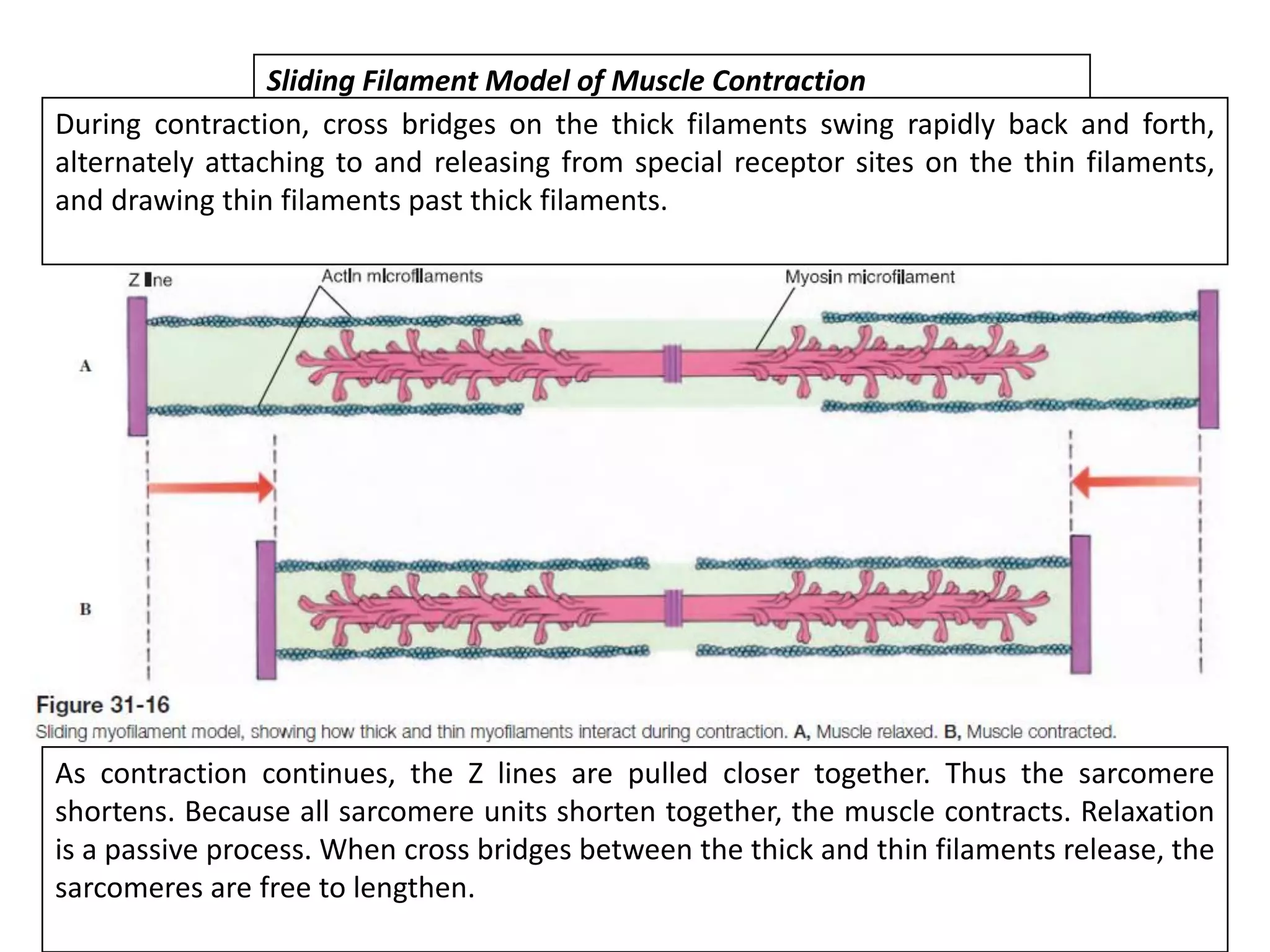
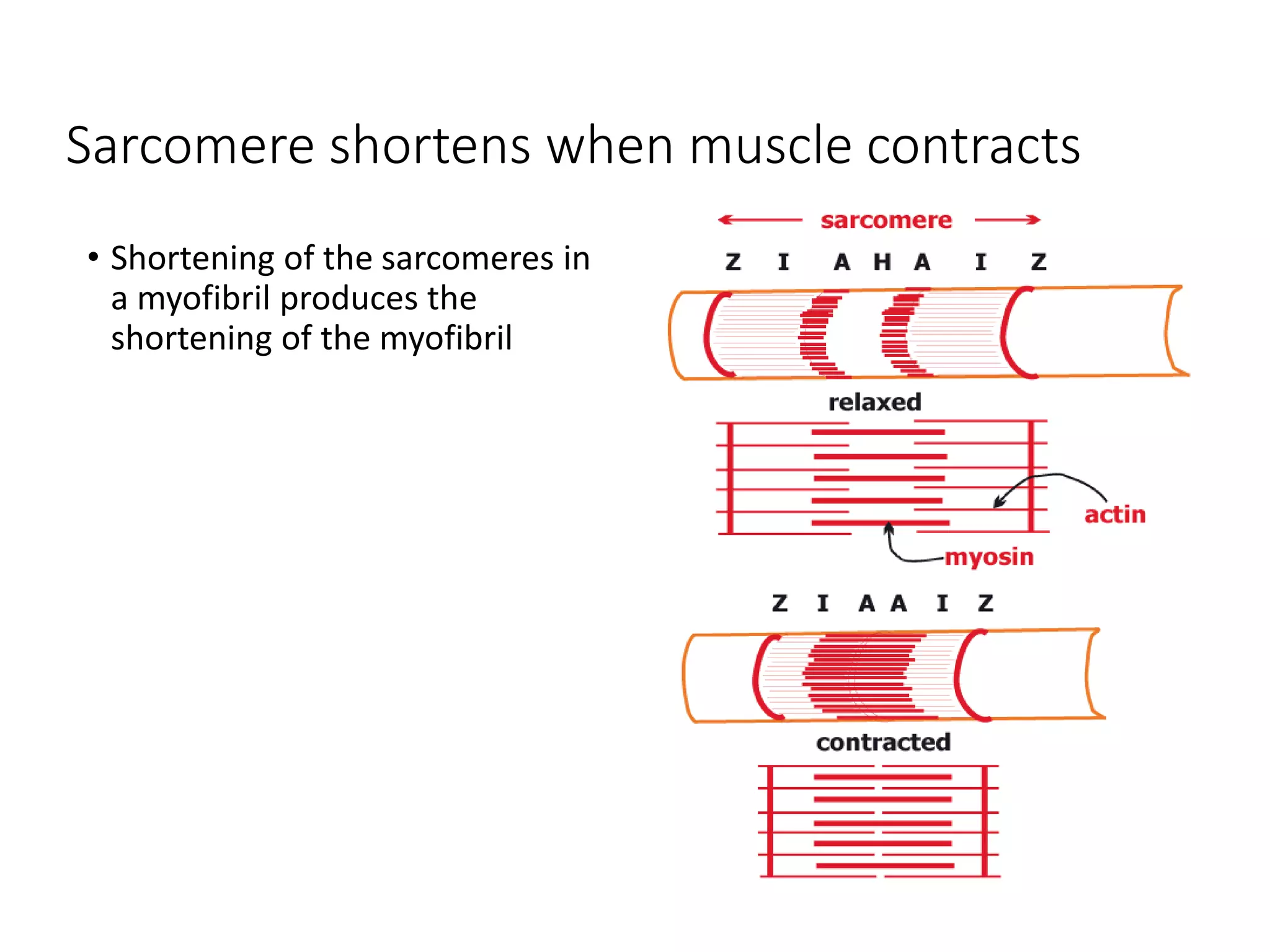
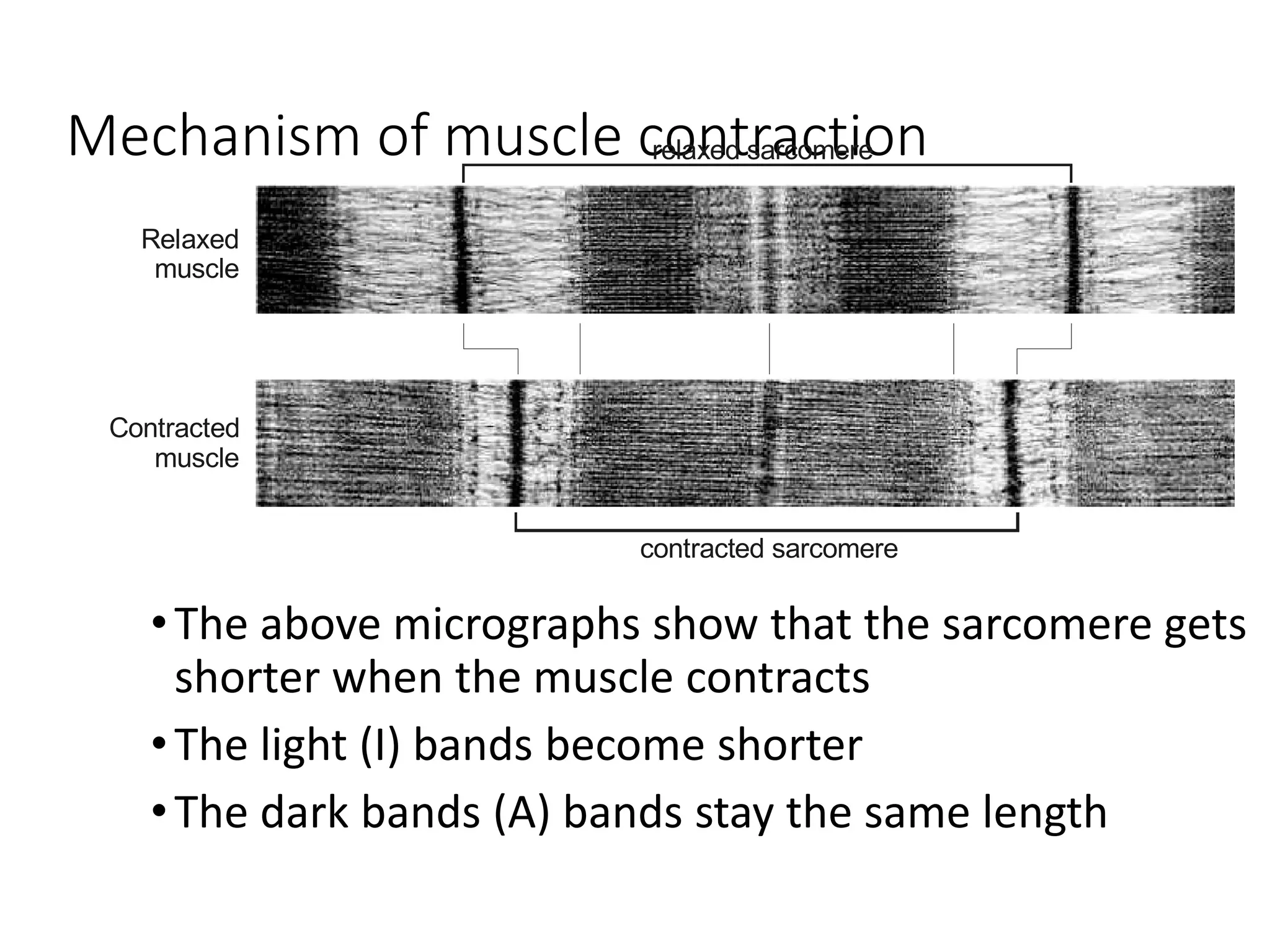


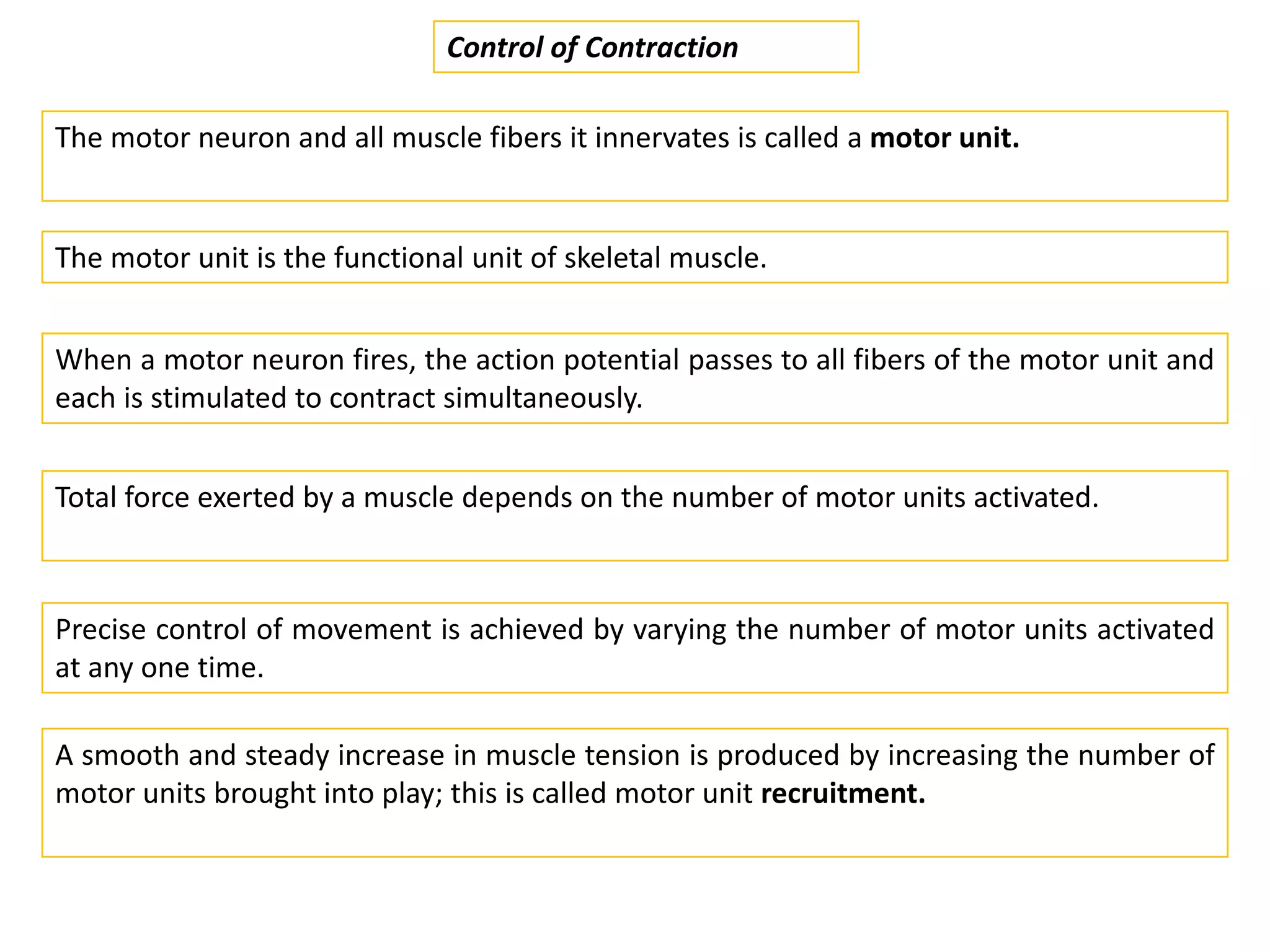
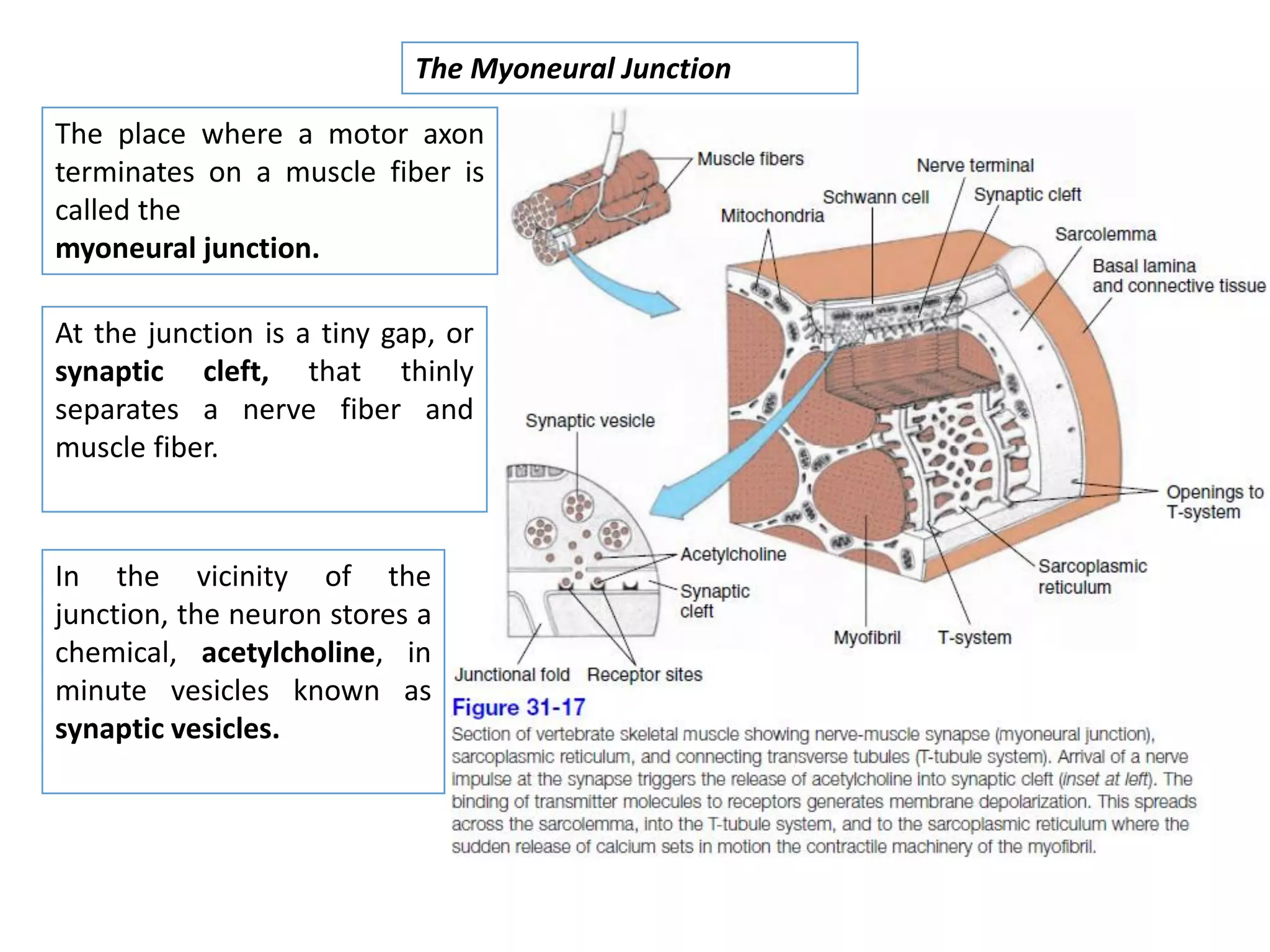
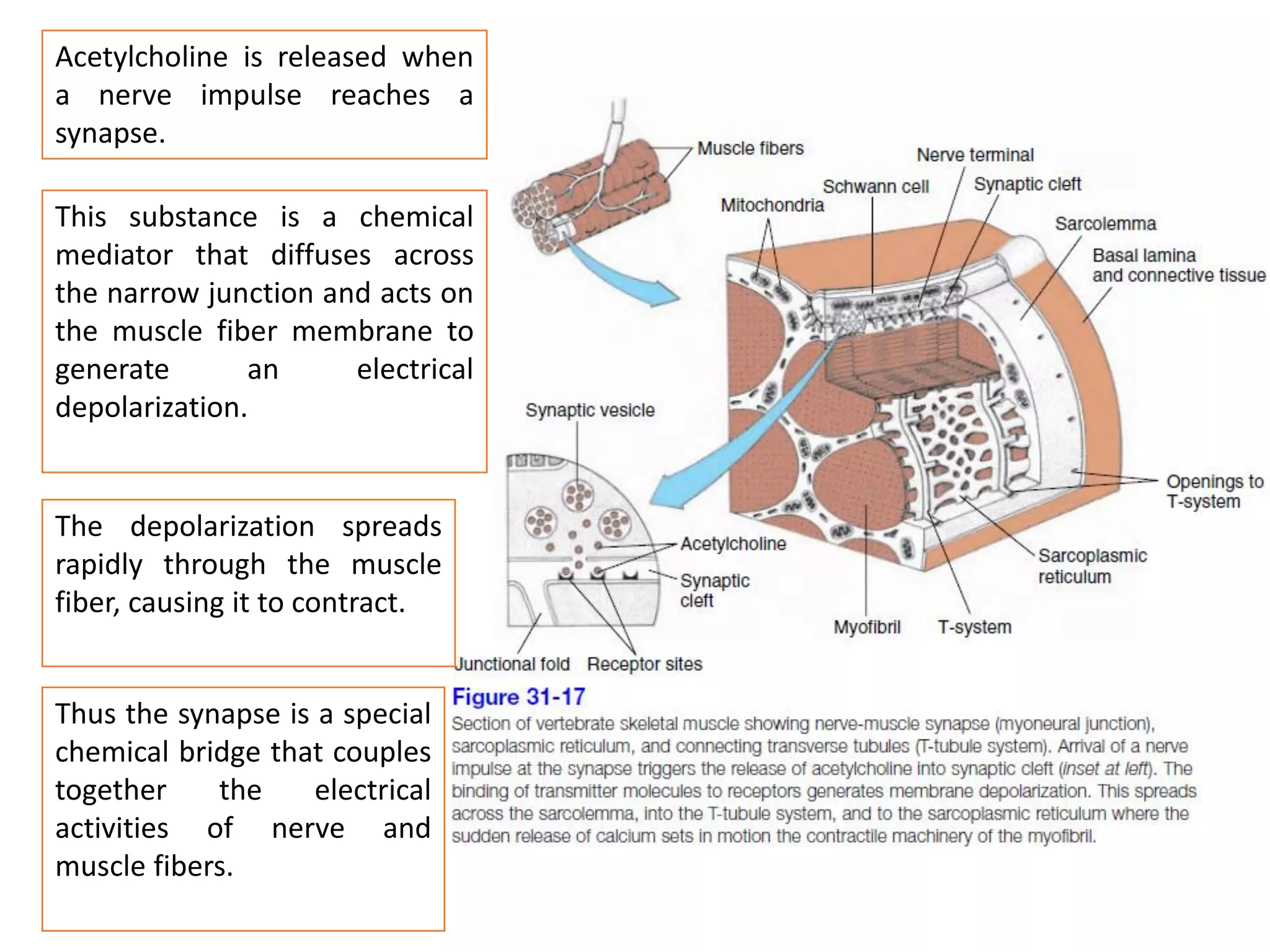
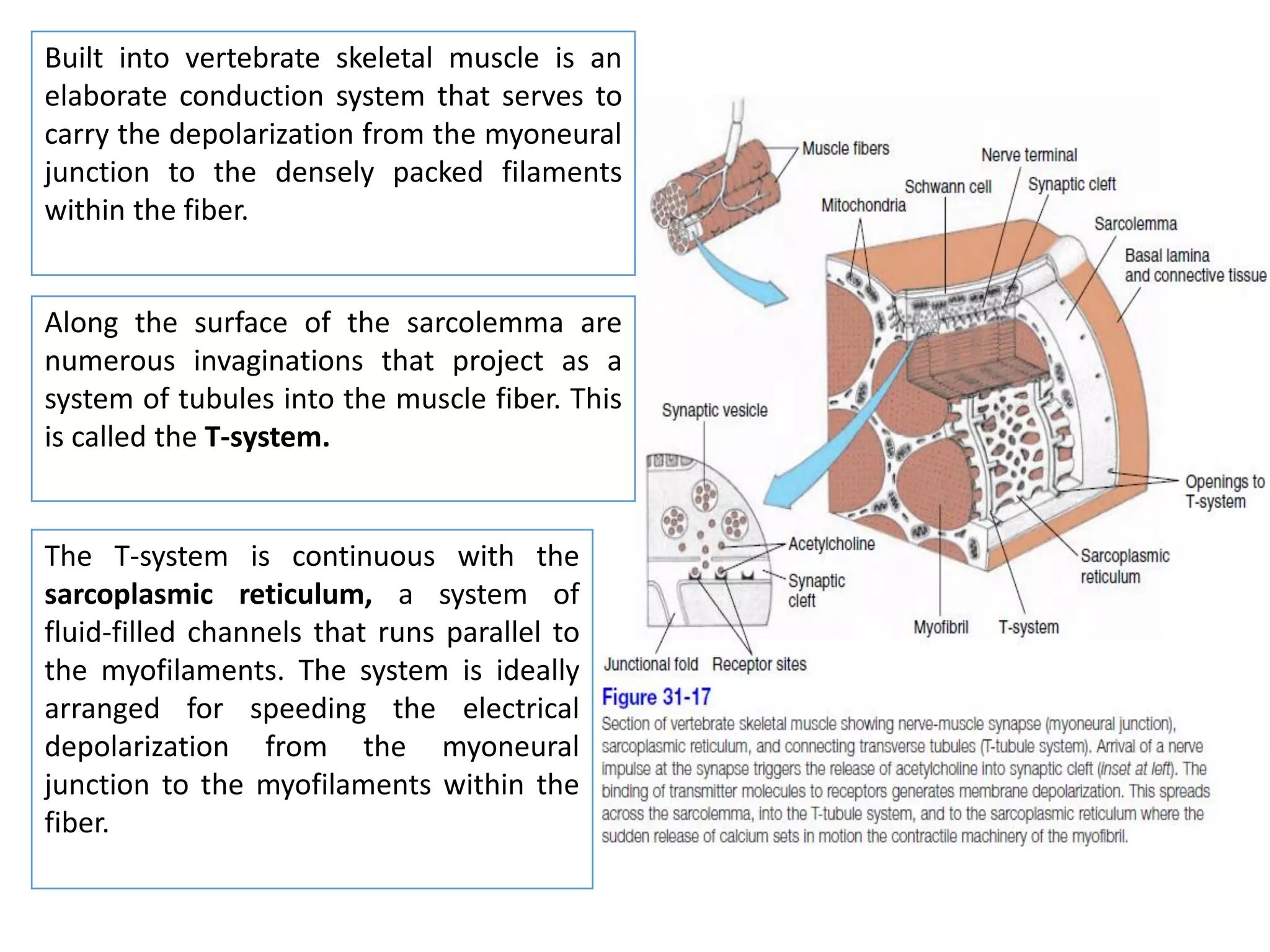
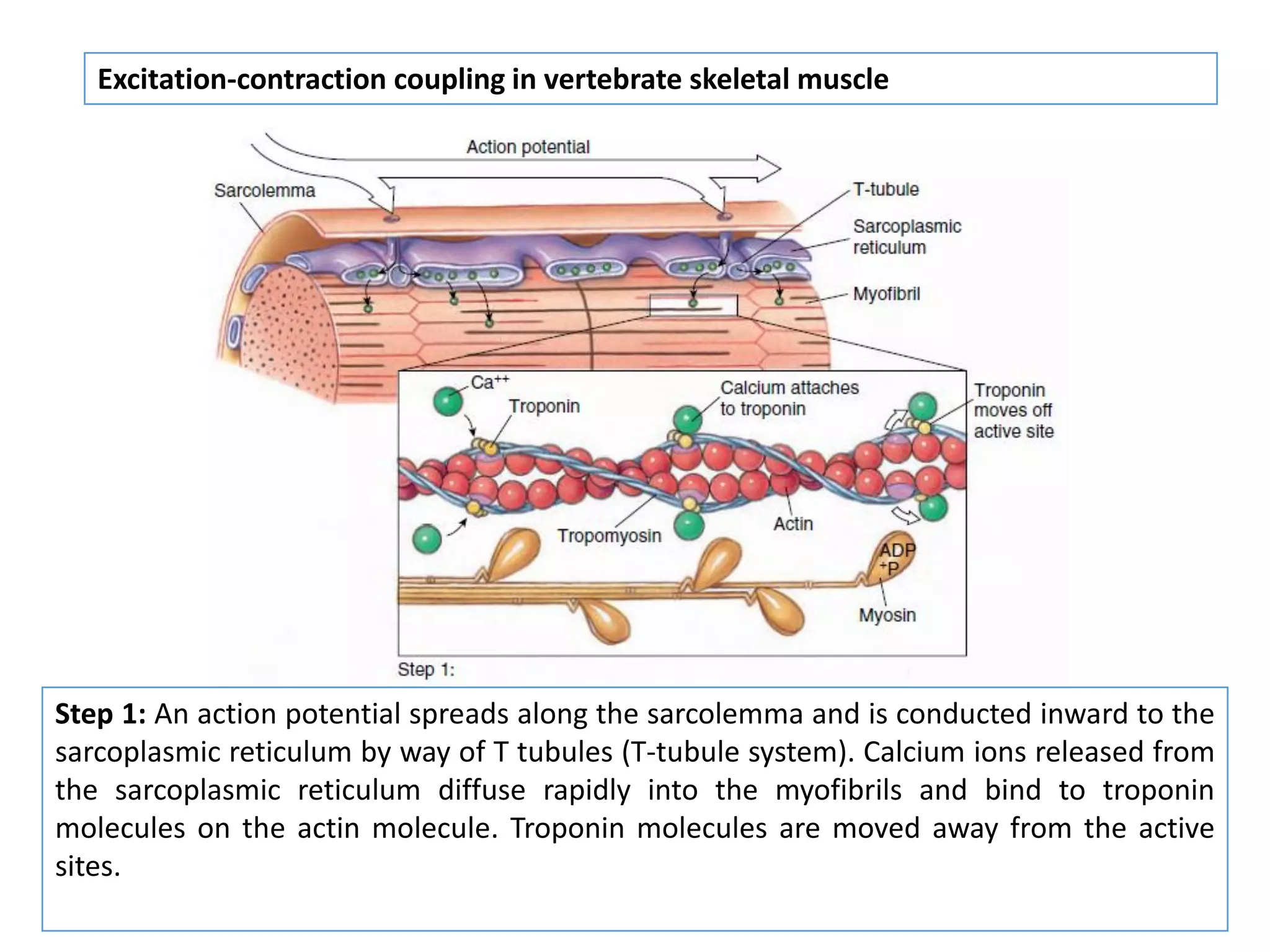
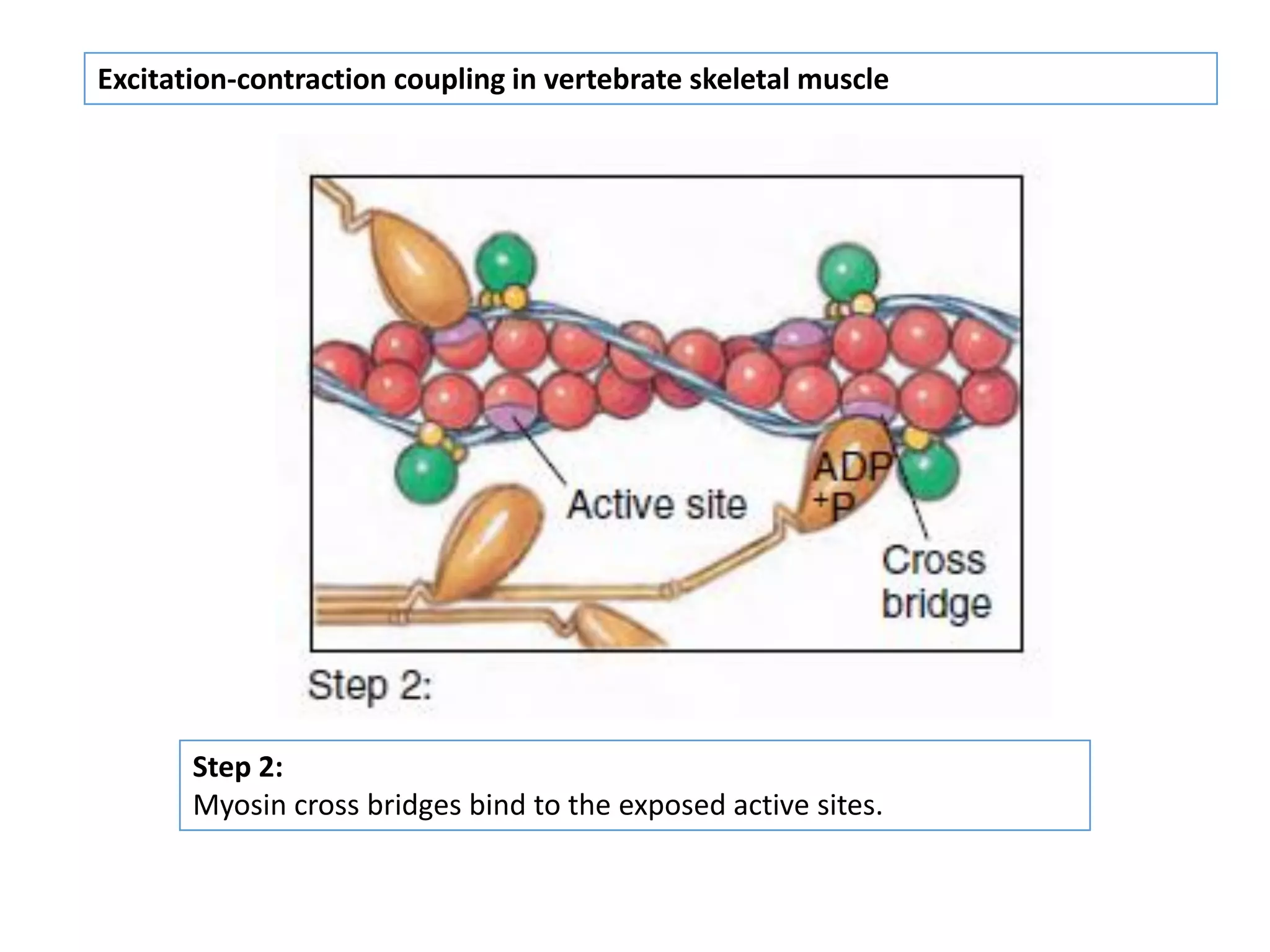
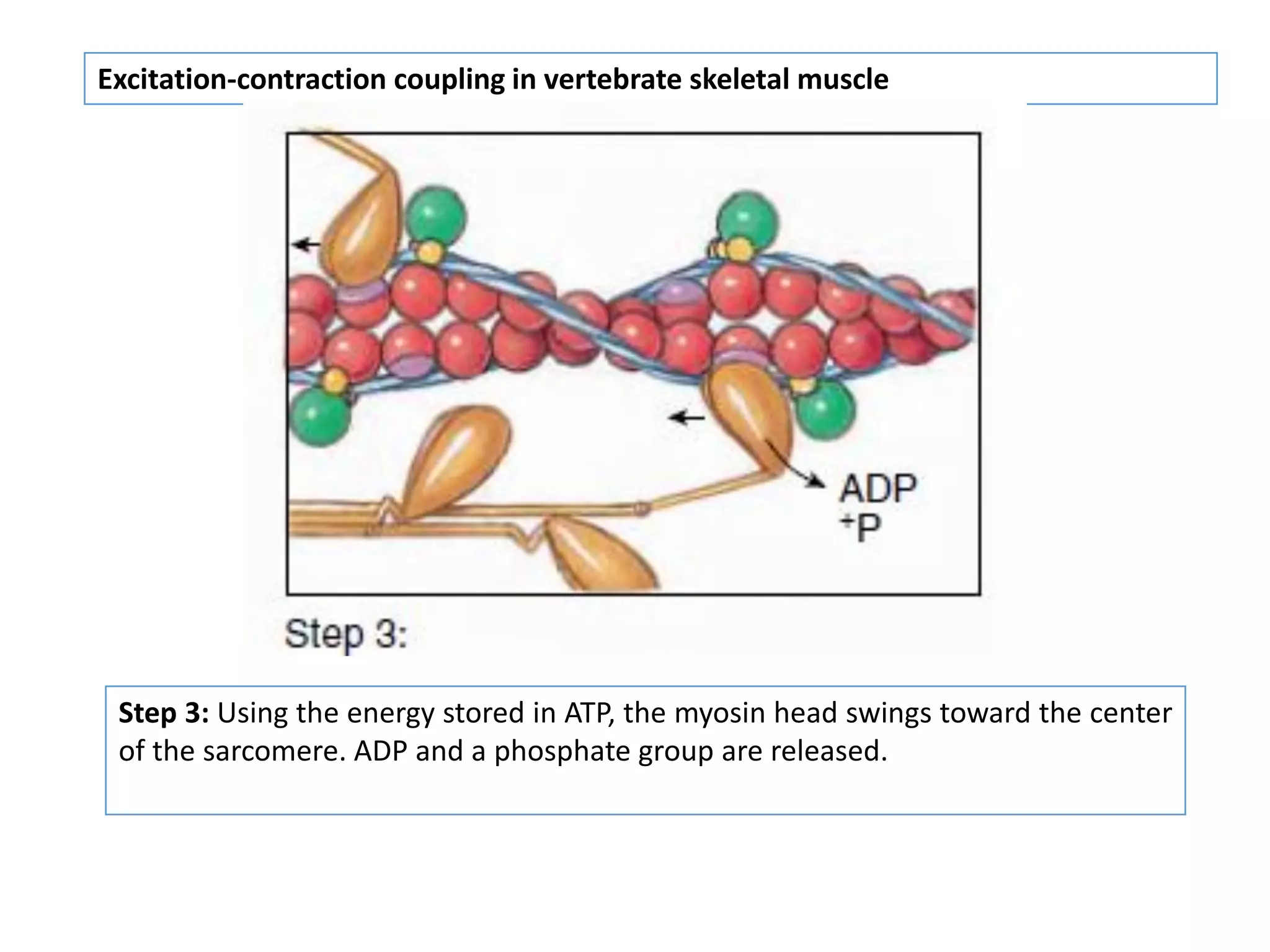
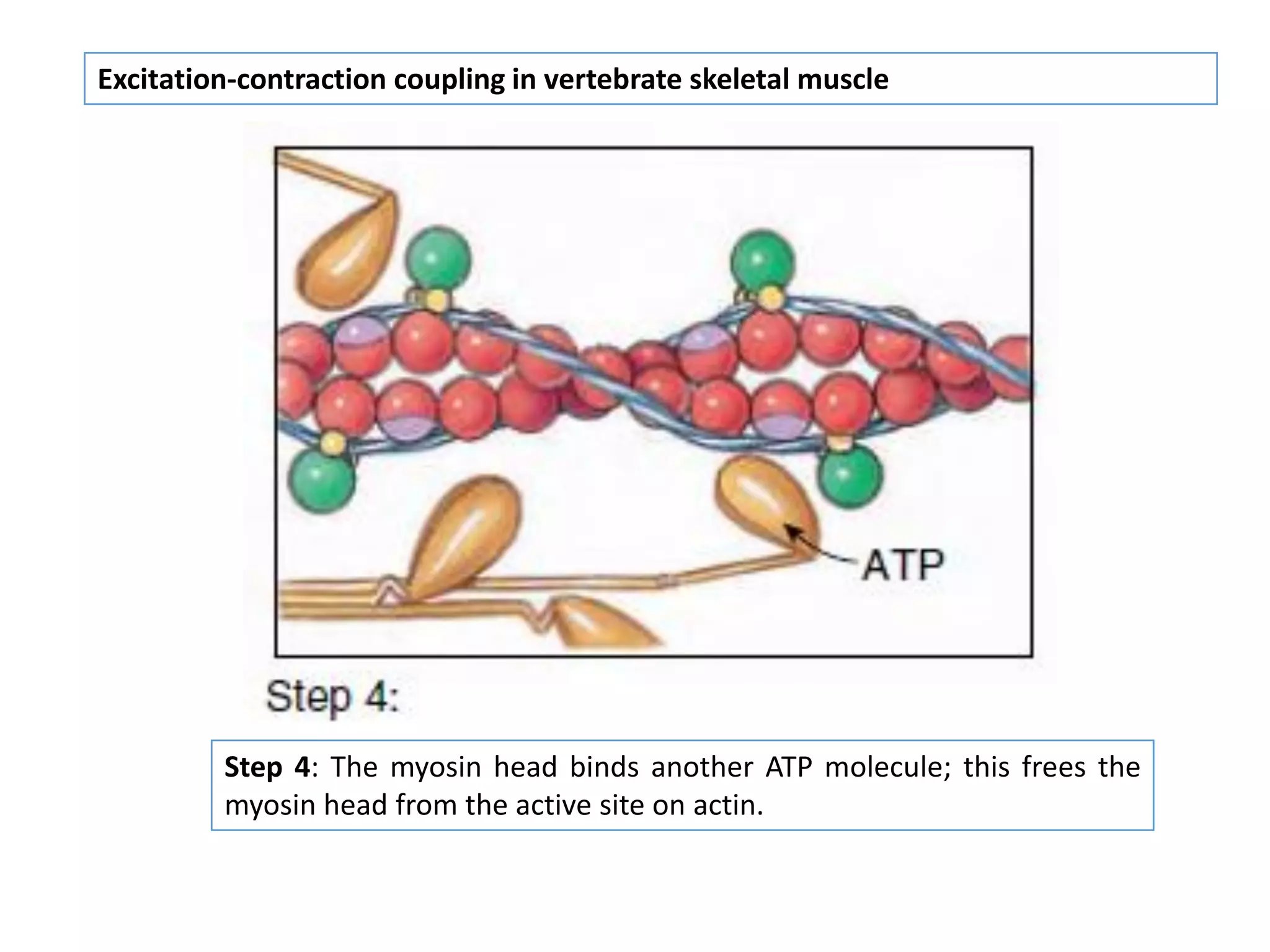
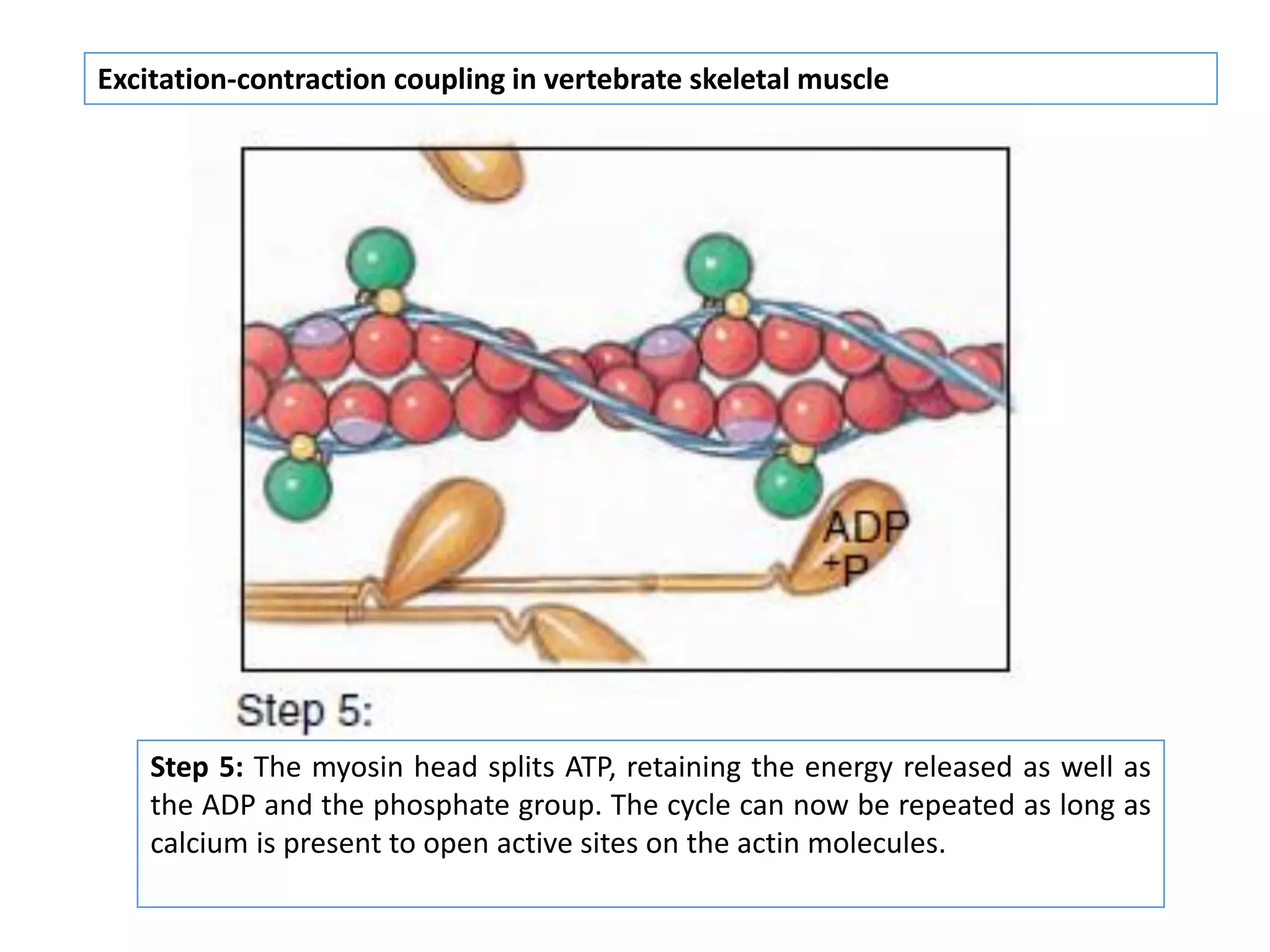

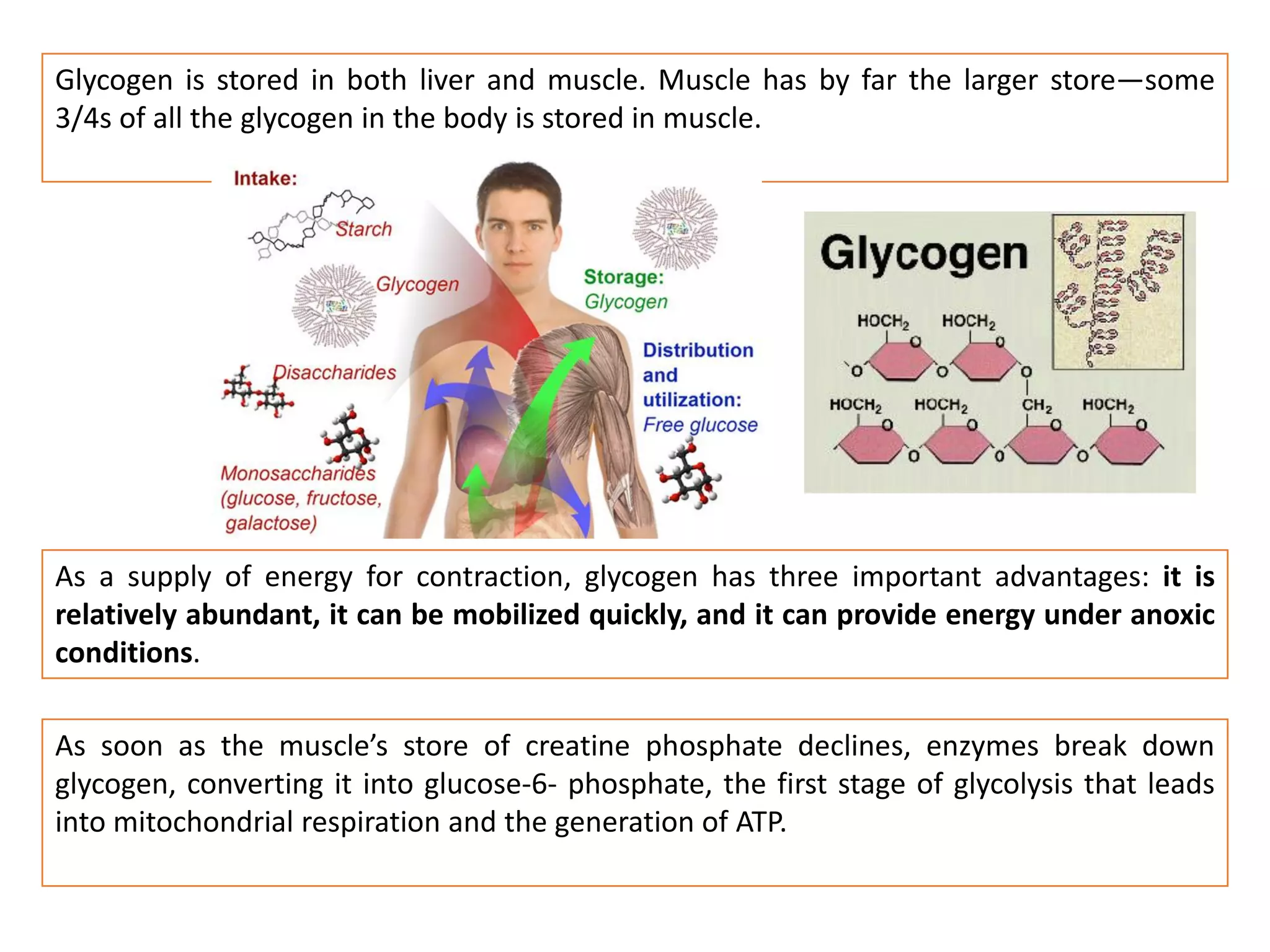
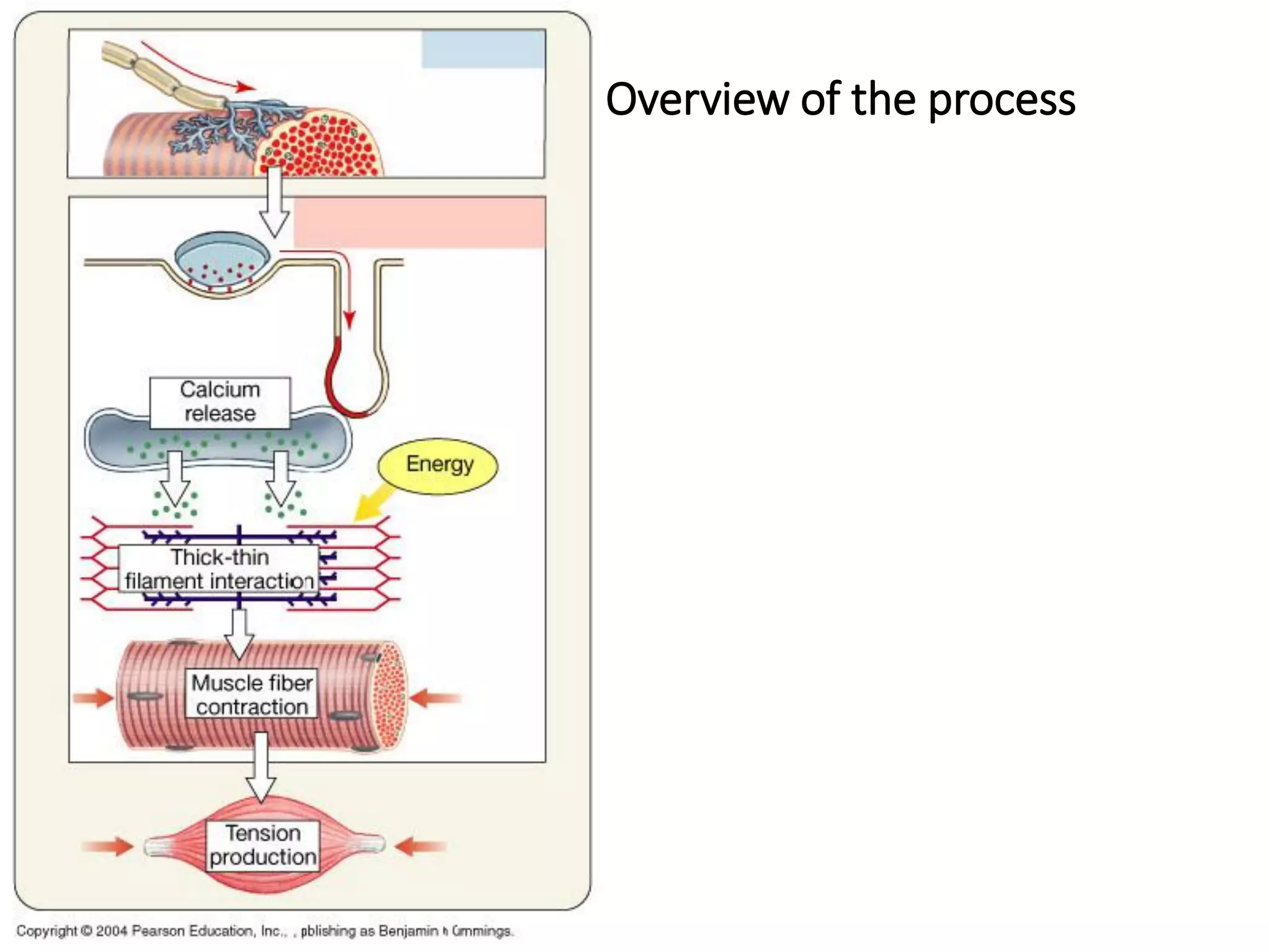
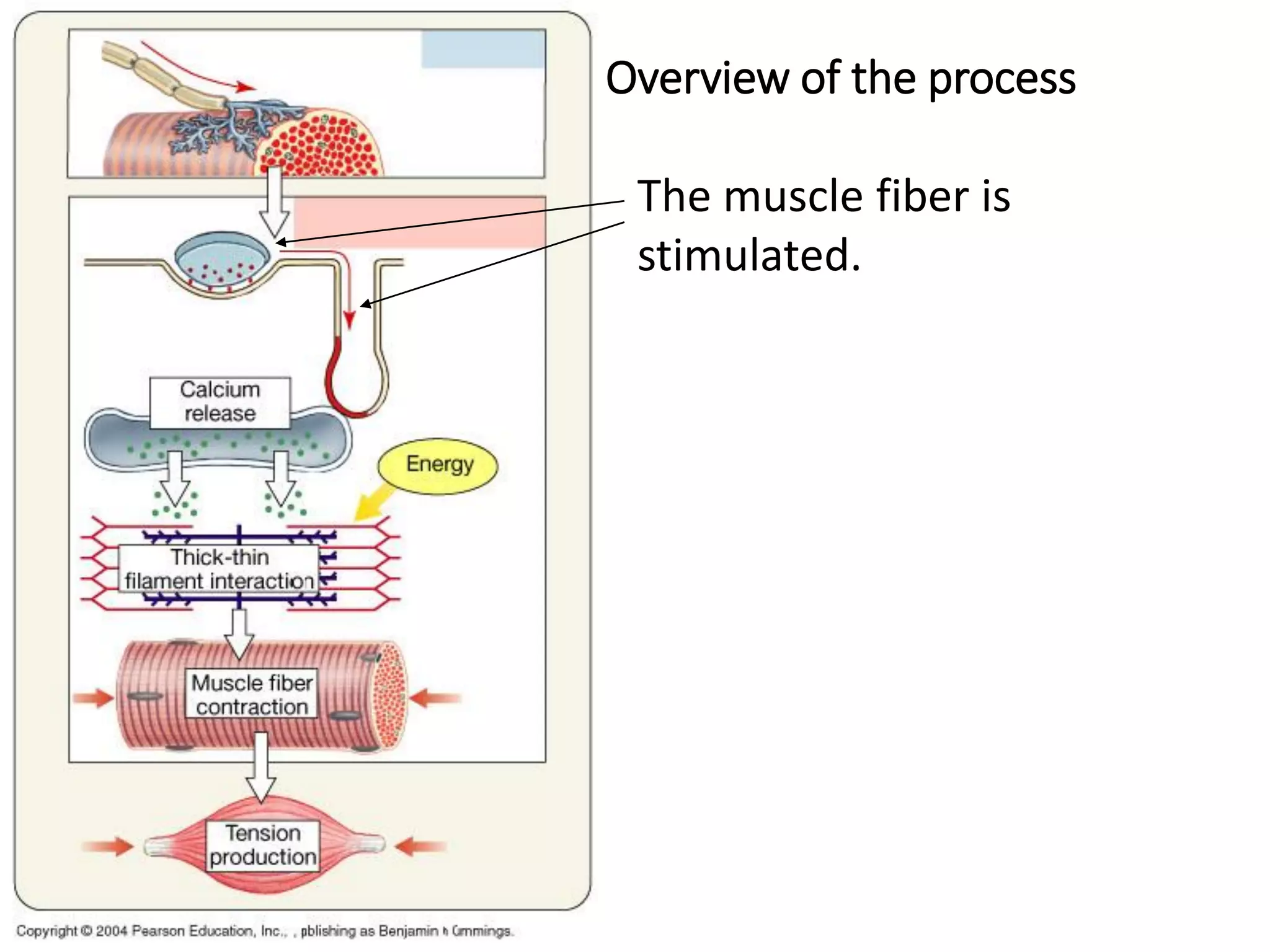
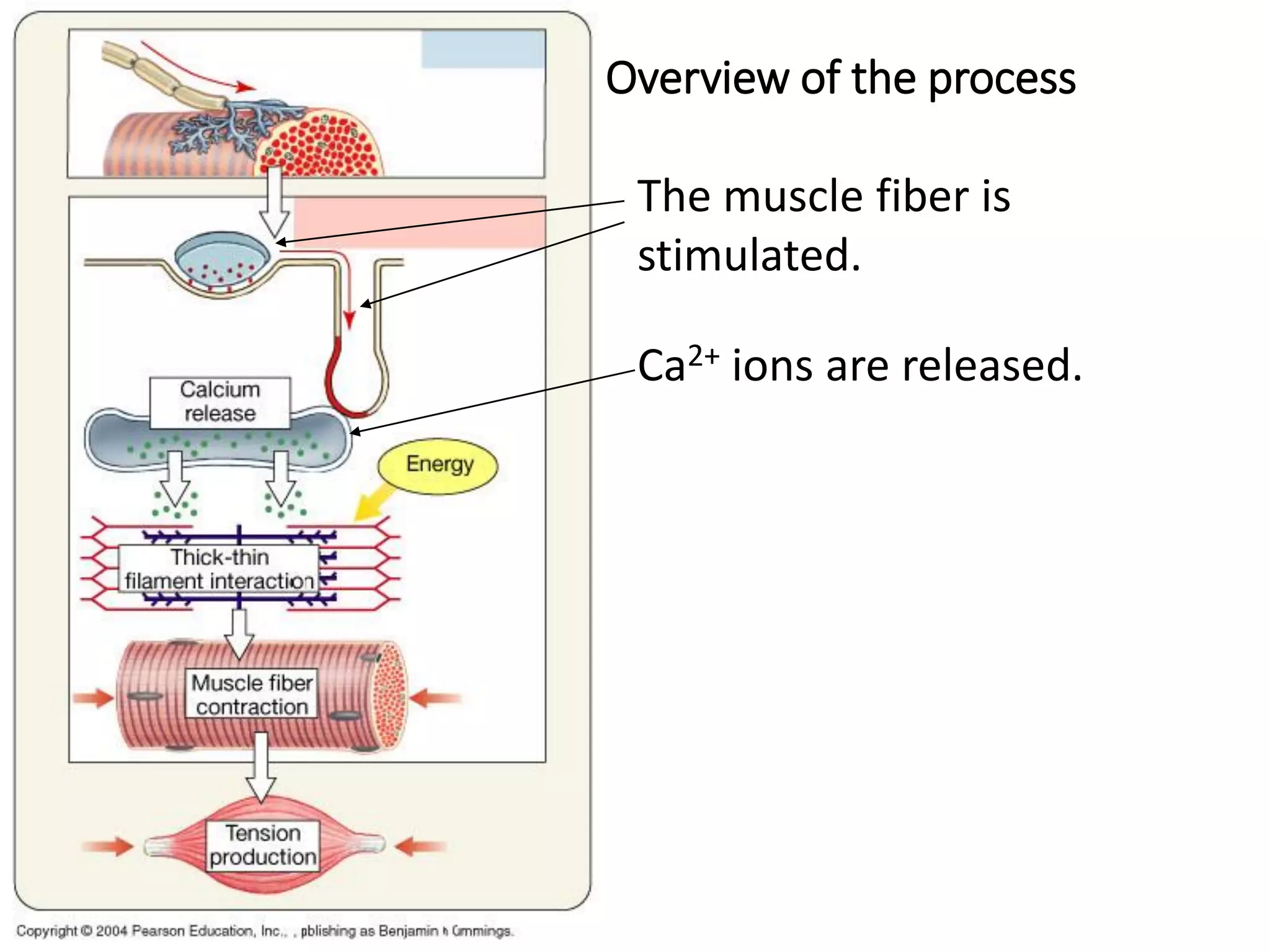
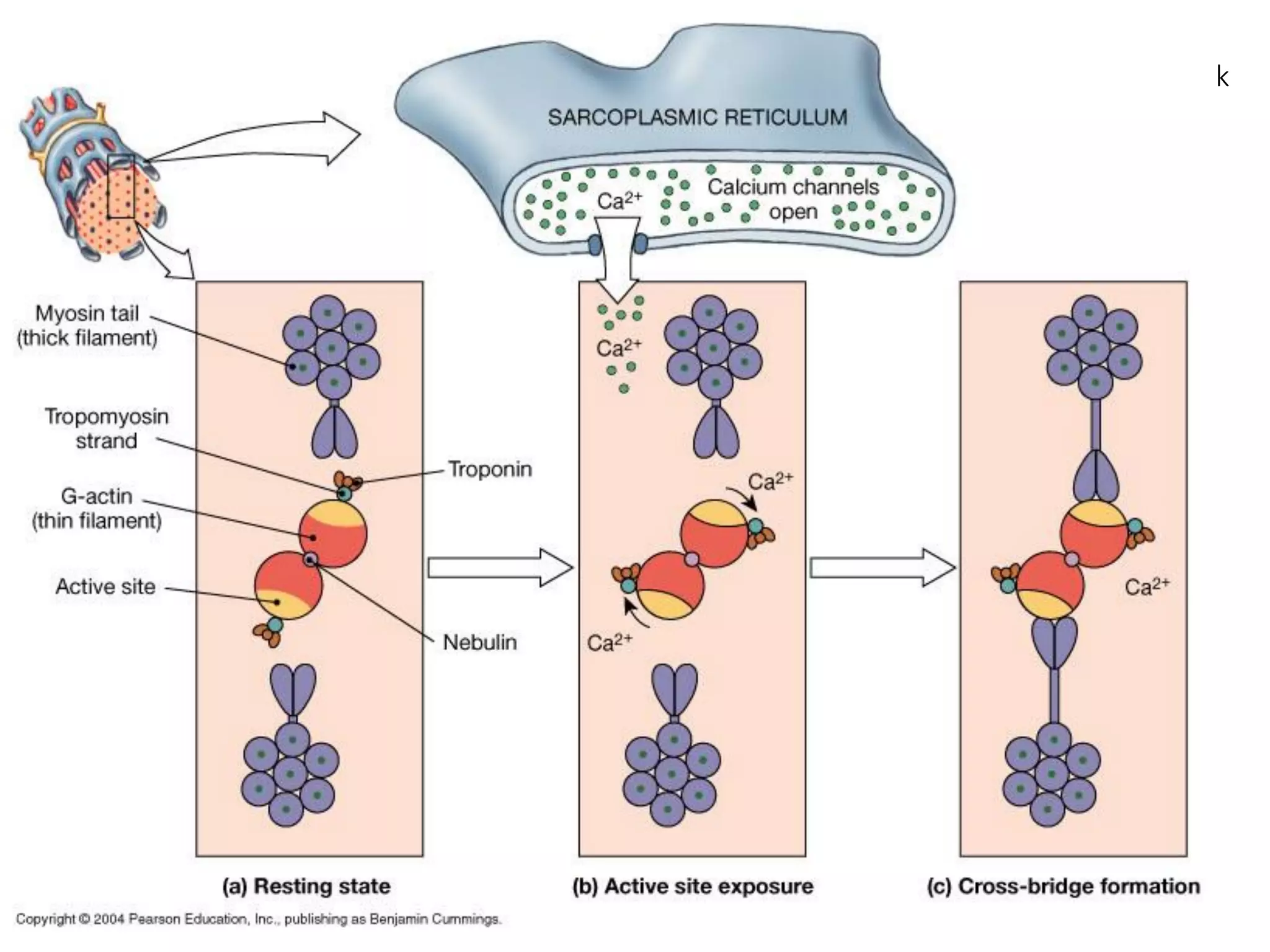
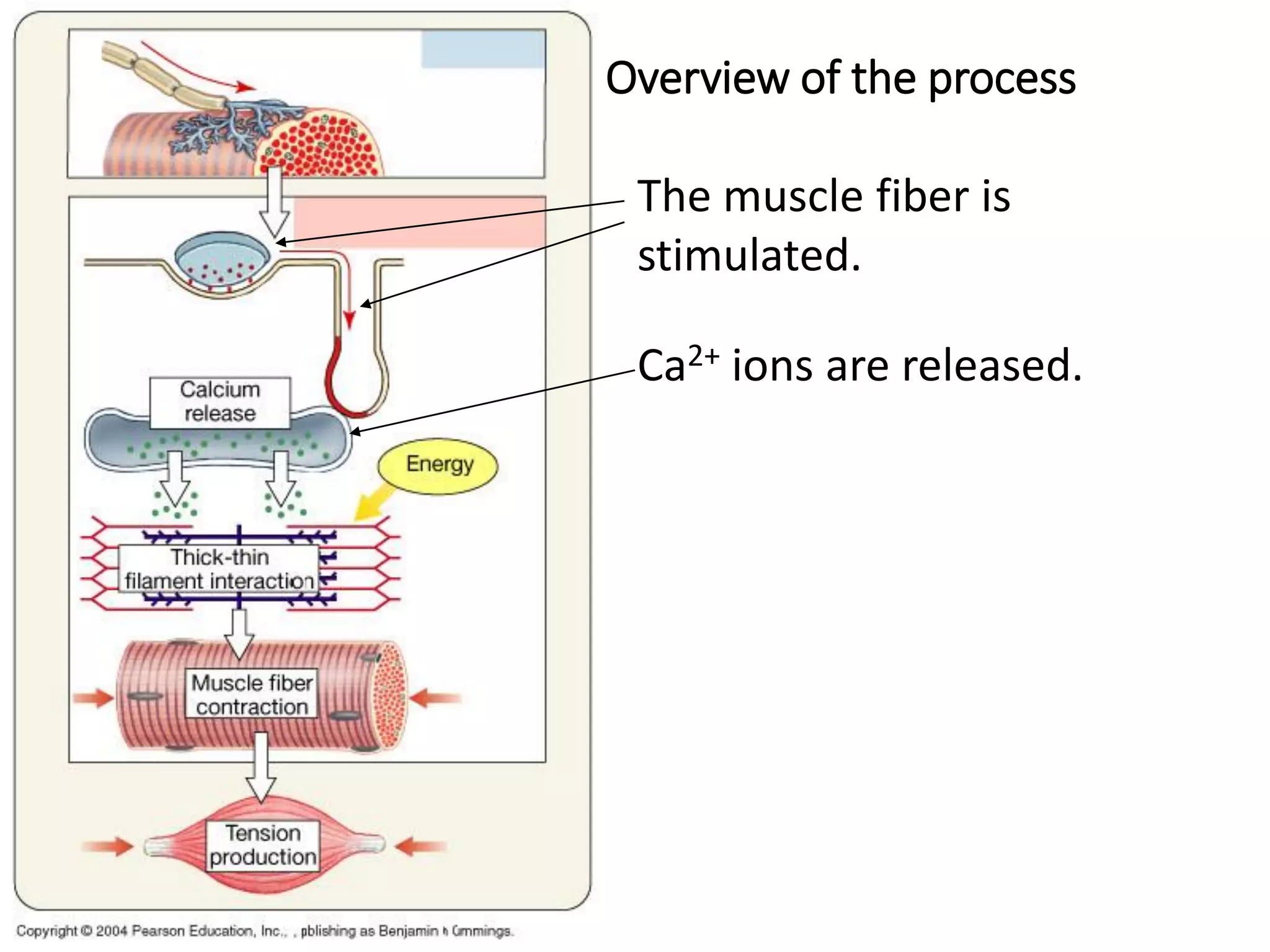
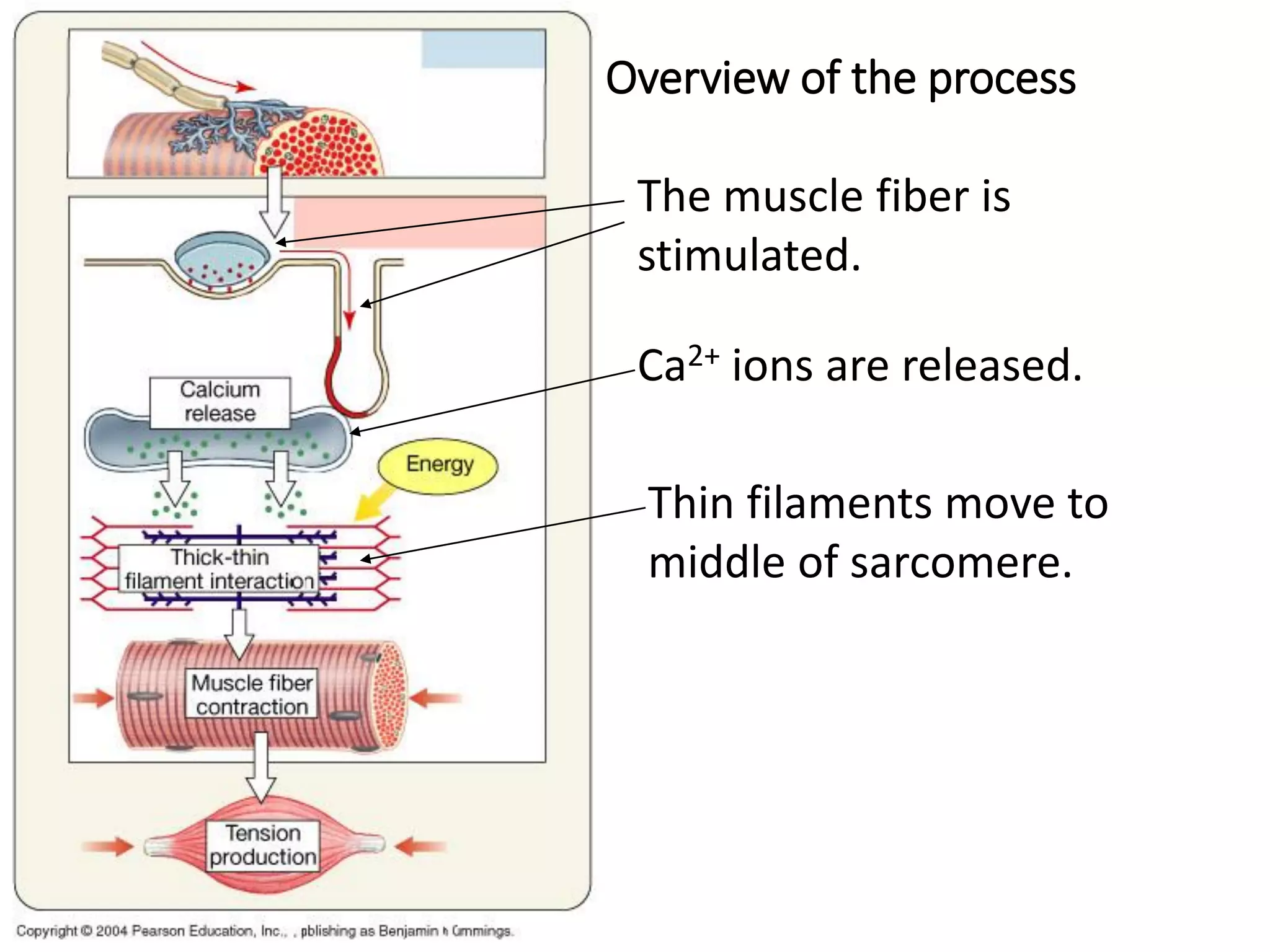
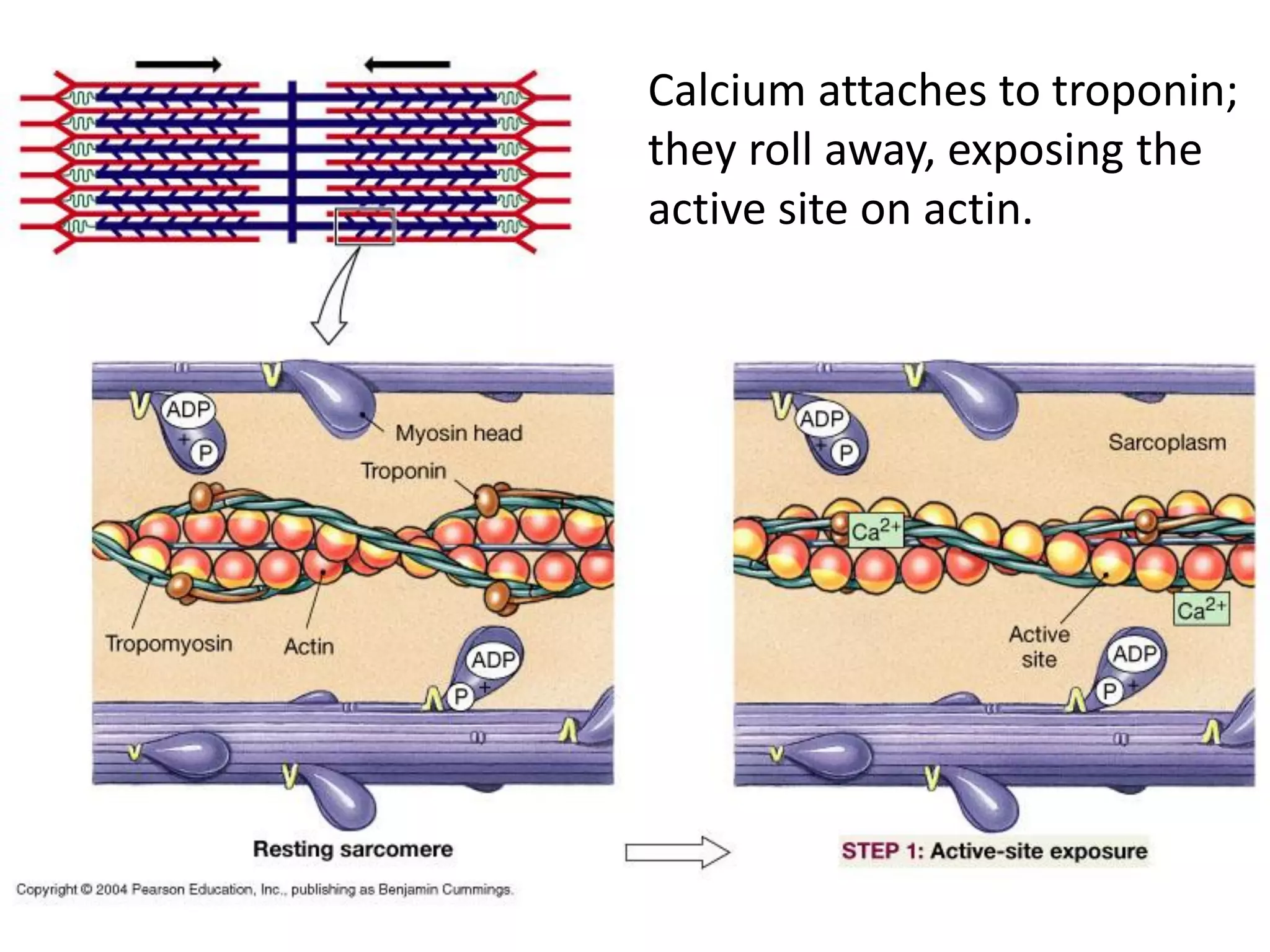
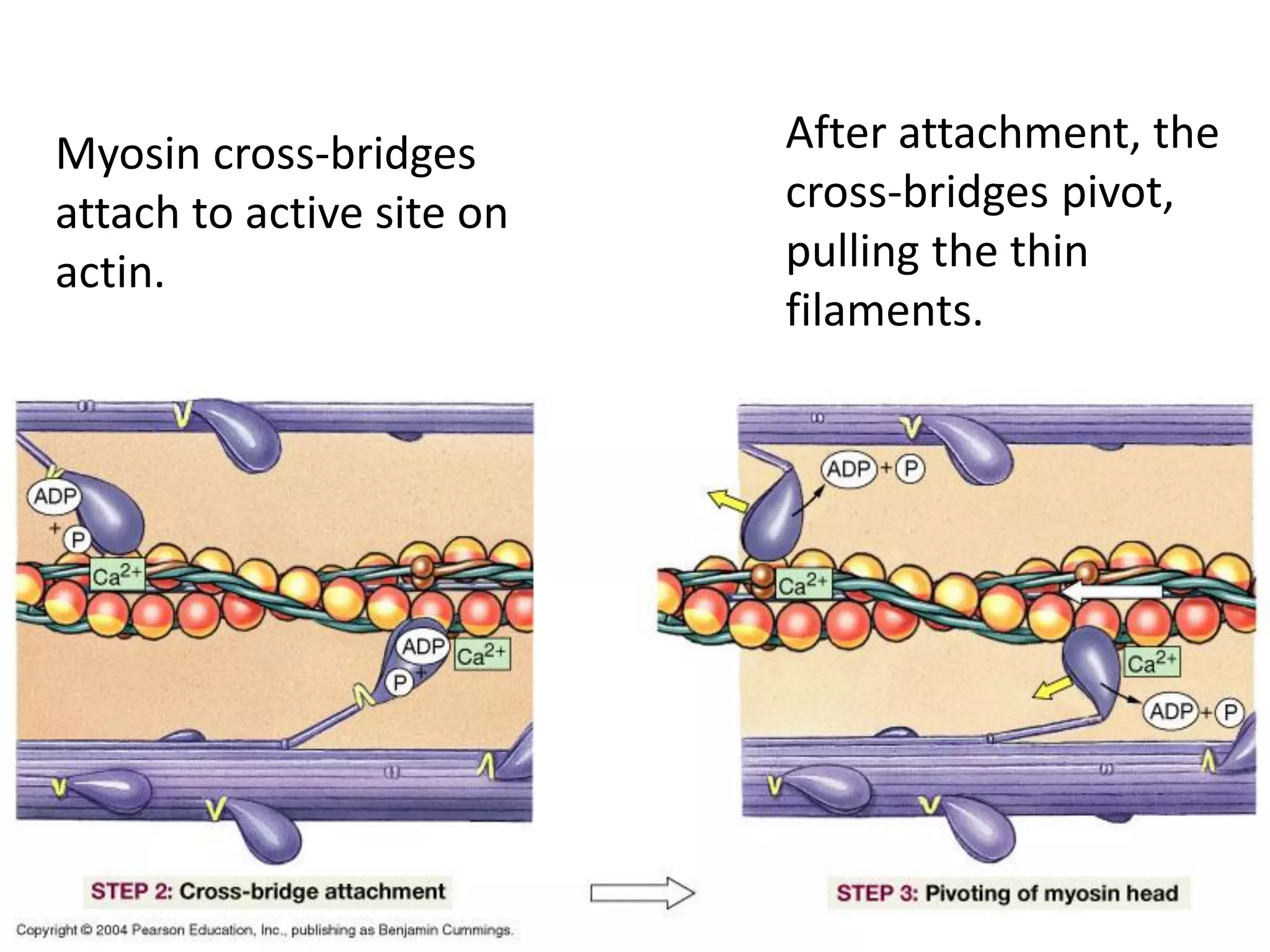
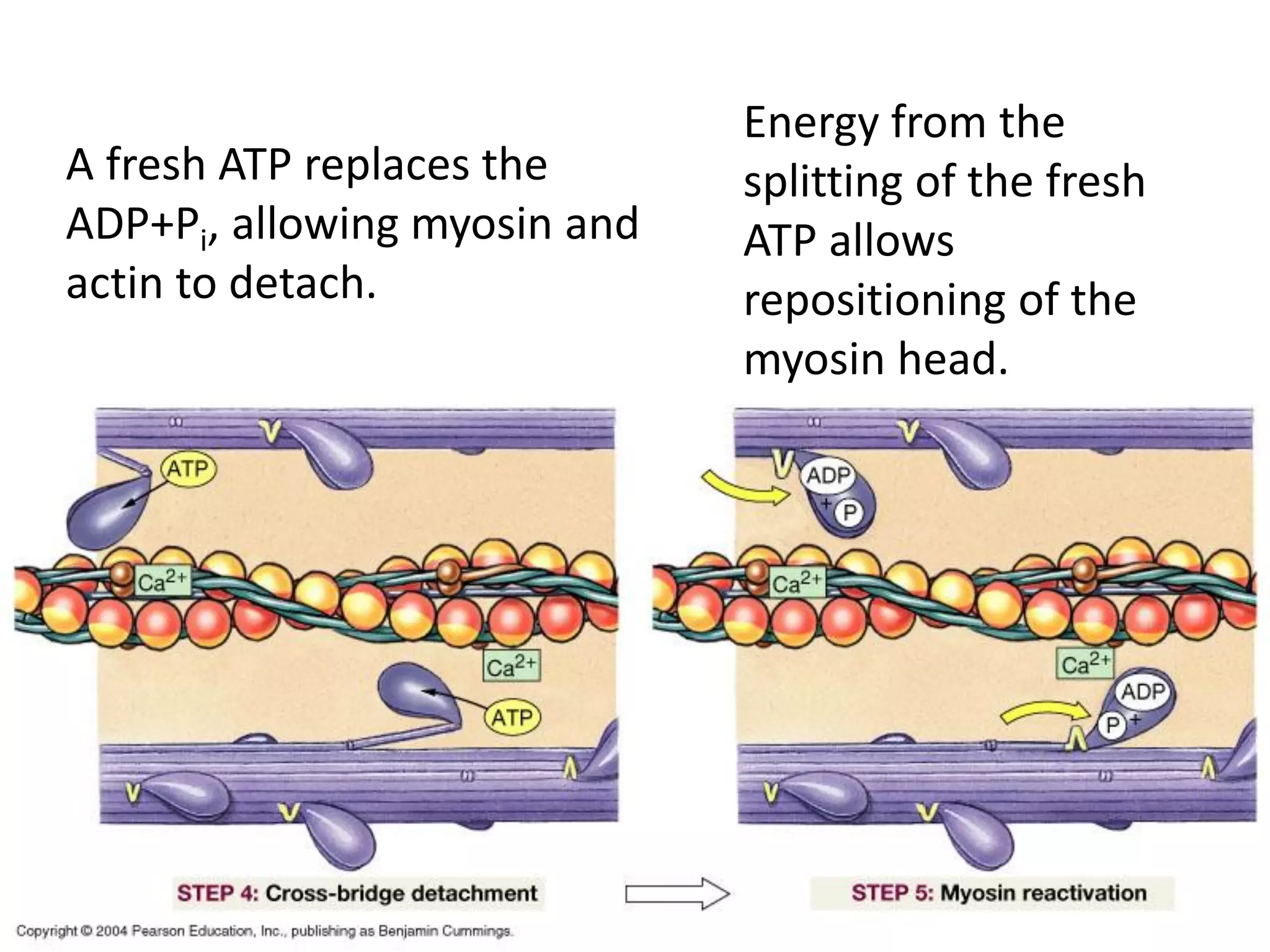
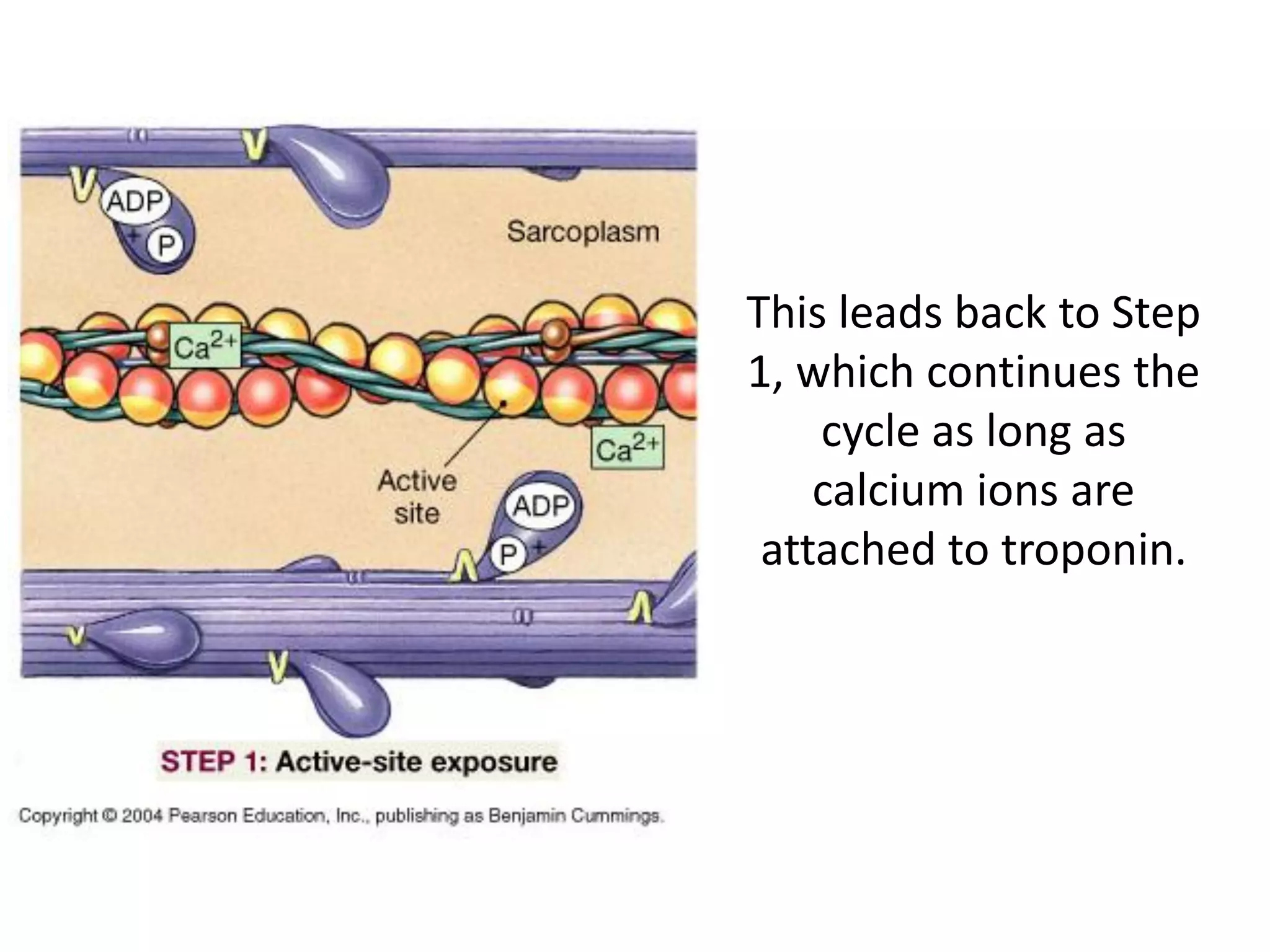
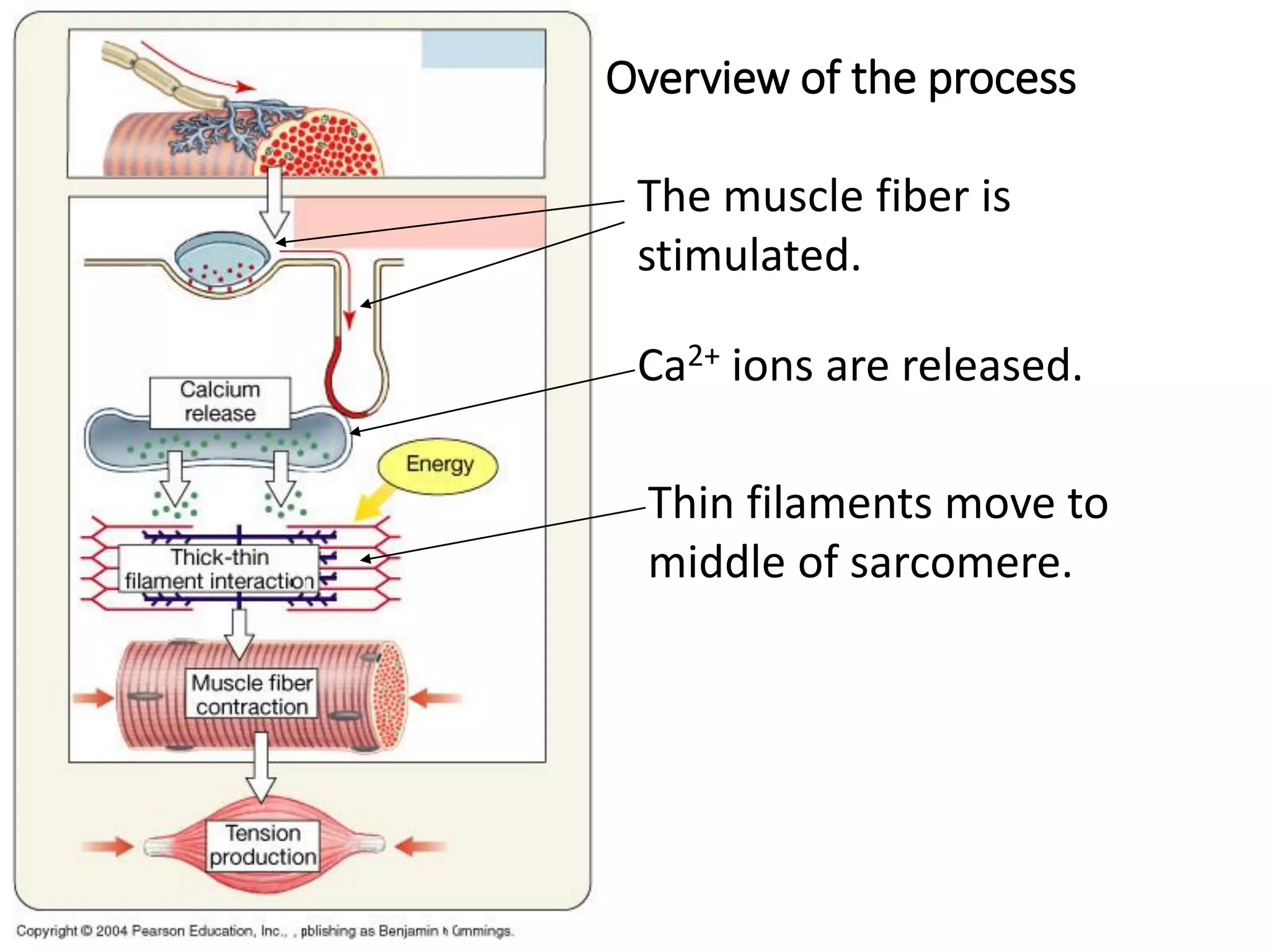
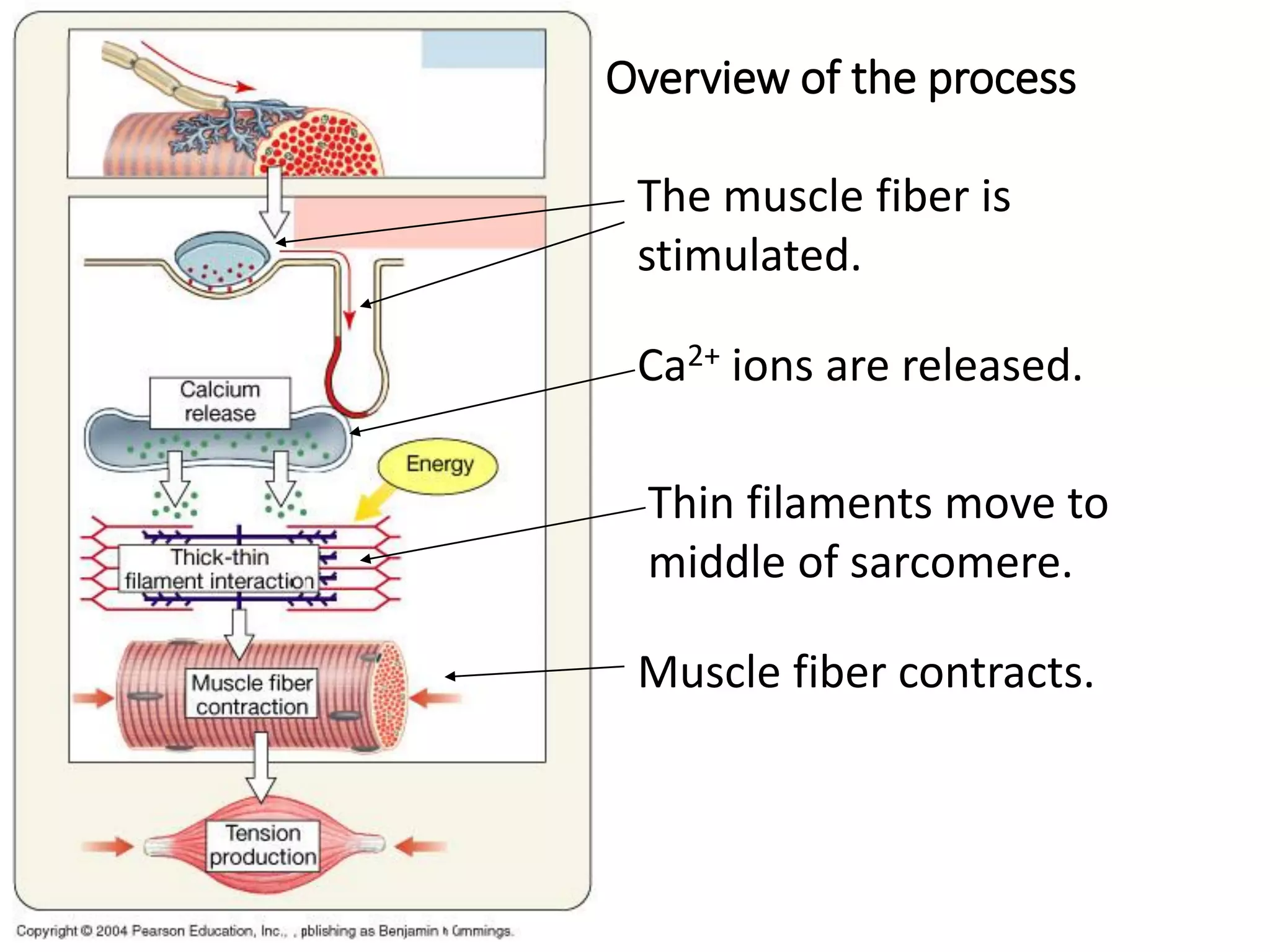
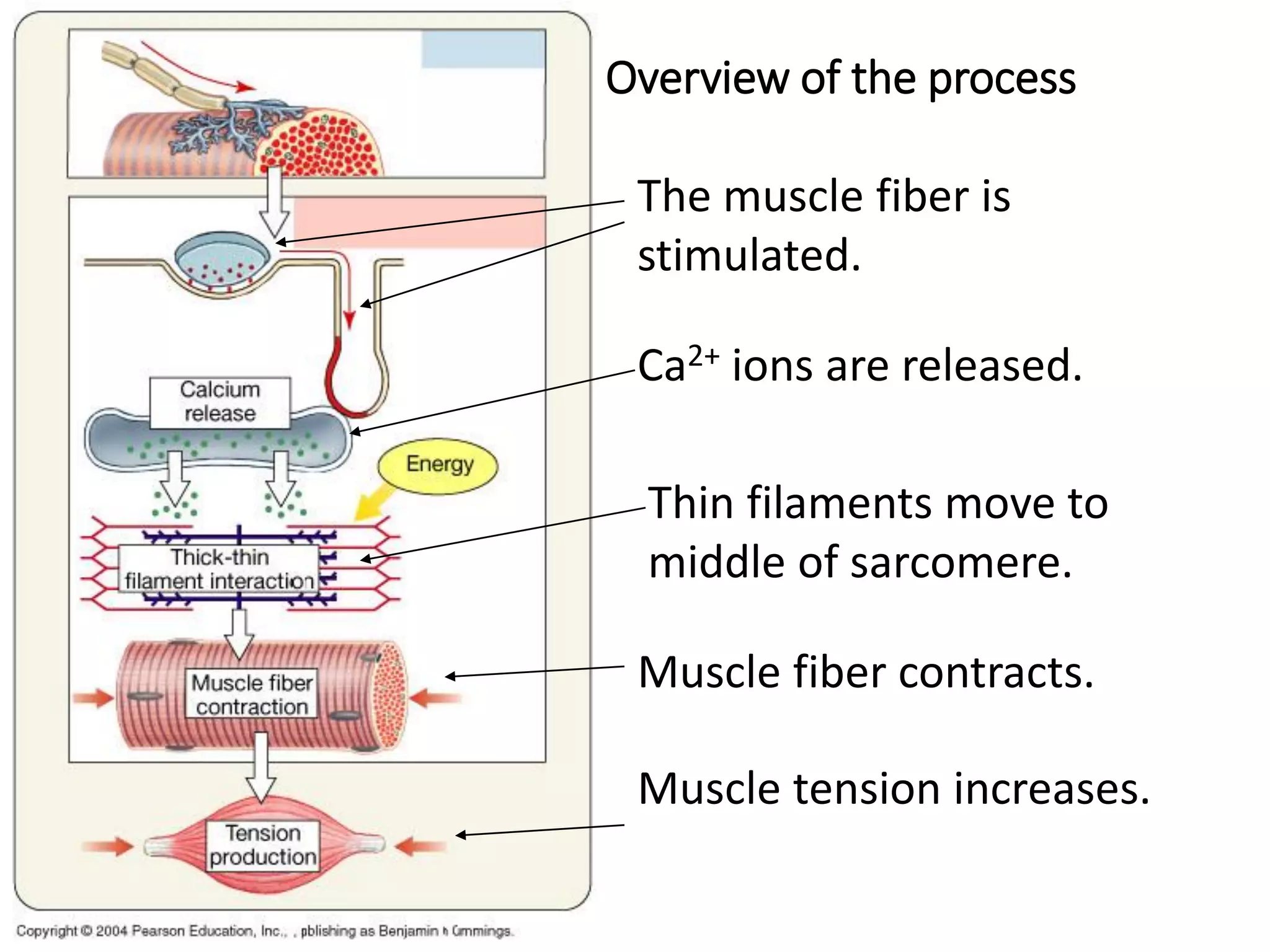



The document summarizes the muscular system and different types of animal movement. There are three main types of movement - ameboid, ciliary, and muscular. Ameboid movement occurs in amebas and other cells via pseudopodia. Ciliary and flagellar movement propel fluids and materials via hairlike cilia and flagella. Muscular movement is powered by the contraction of muscle fibers and occurs in three main types of muscle - skeletal, smooth, and cardiac. Skeletal muscle is striated and voluntary, smooth muscle is involuntary, and cardiac muscle is striated and involuntary. Contraction occurs via the sliding filament model where actin and myosin filaments slide past each other through cross-brid











































































































