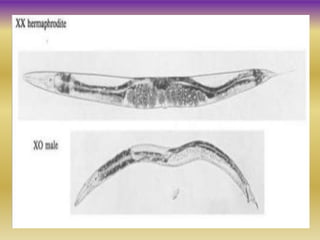
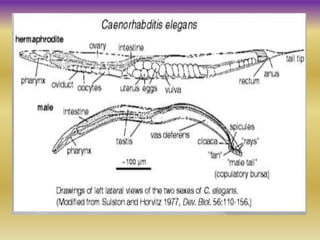
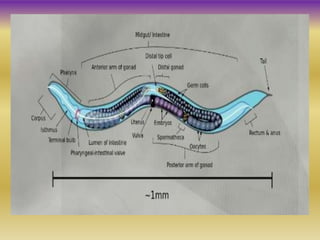
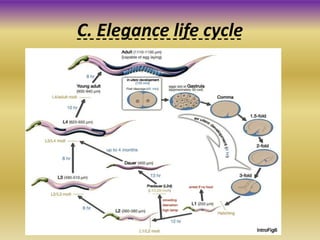
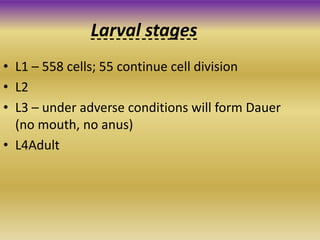
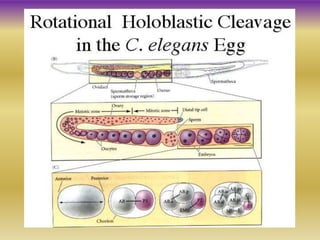
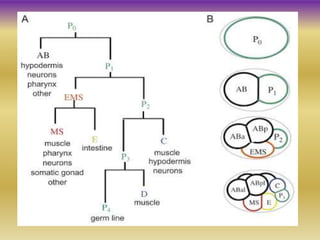
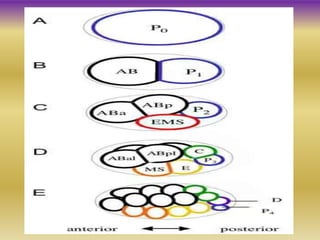
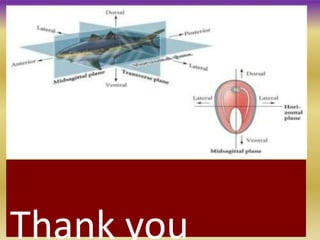
This document summarizes key aspects of the nematode Caenorhabditis elegans. It describes C. elegans' structure as a 1mm long roundworm with 945 cells. It details its life cycle from zygote to adult over 3 days, producing 300 progeny. C. elegans establishes three embryonic axes - anterior-posterior determined by sperm entry point, dorso-ventral set by first cleavage, and left-right by asymmetric cell divisions. The document outlines C. elegans' larval stages and early embryonic development, including axis formation and cell lineage.