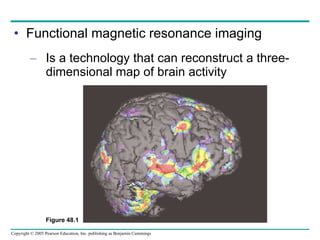
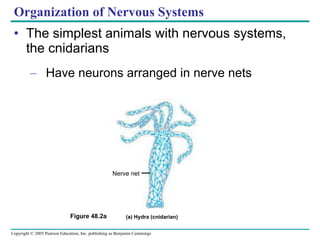
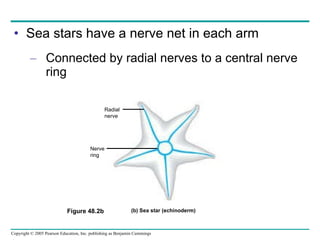
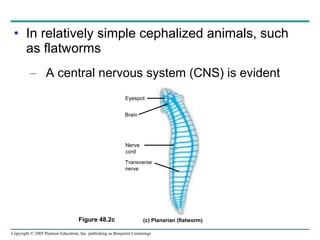
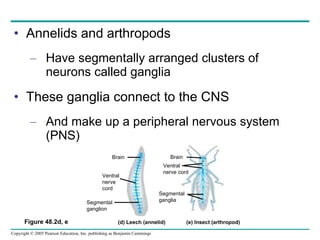
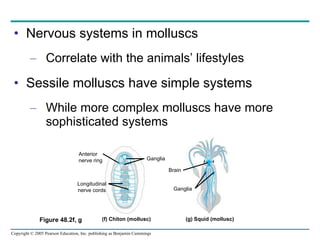
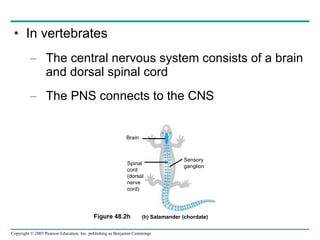
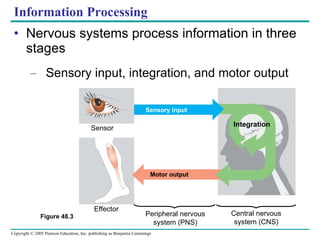
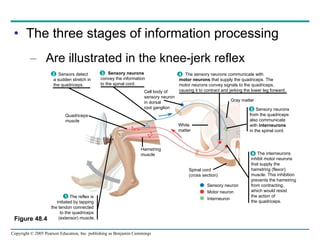
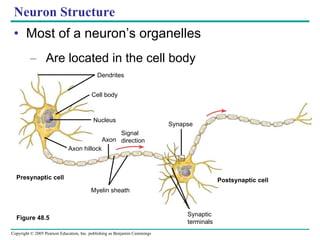
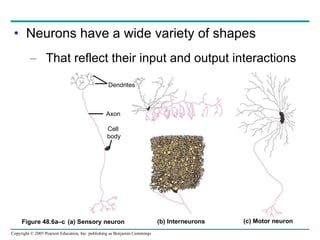
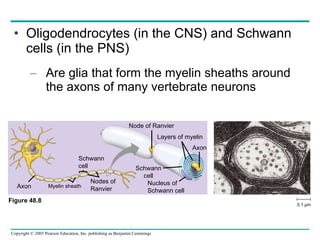
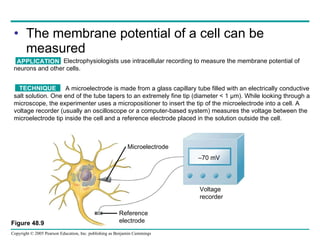
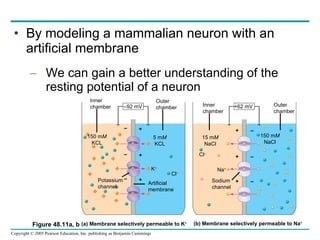
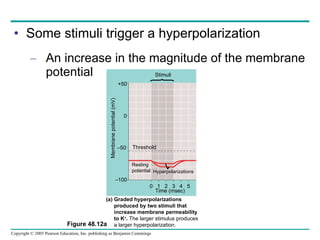
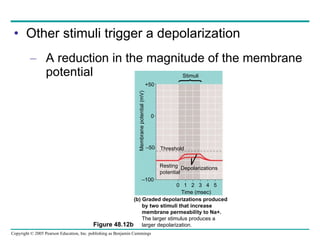
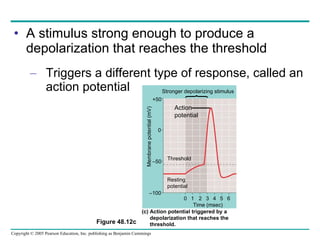
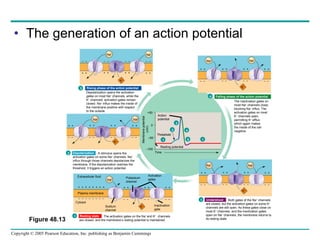
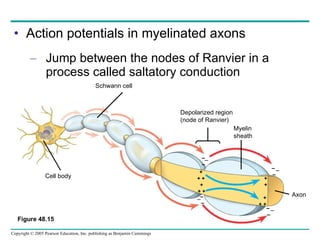
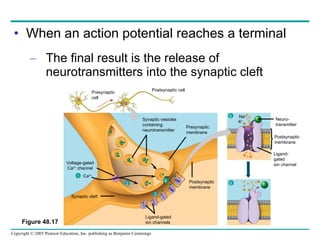
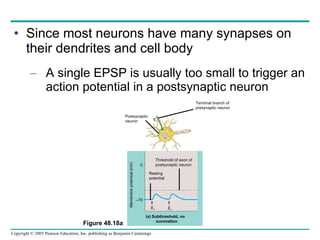
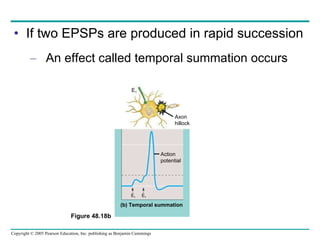
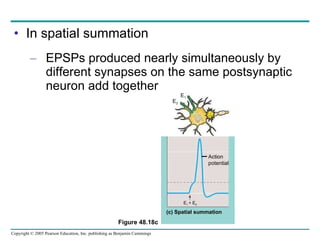
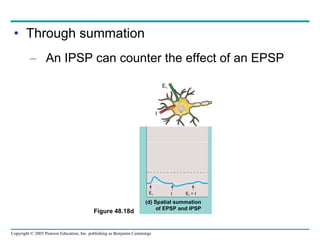
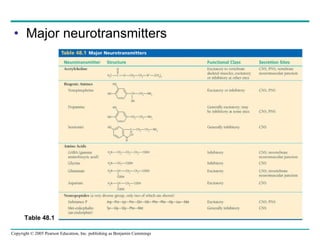
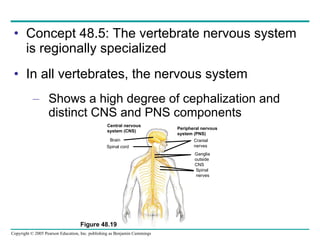
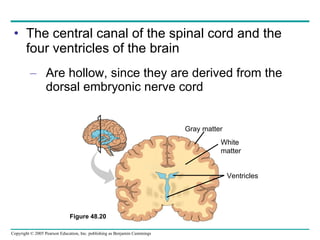
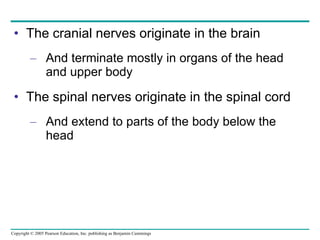
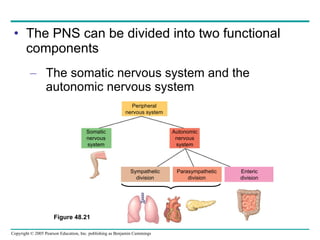
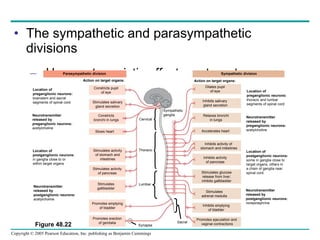
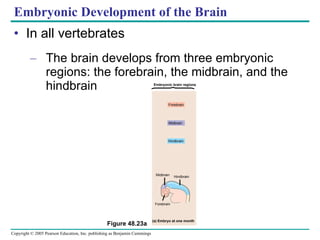
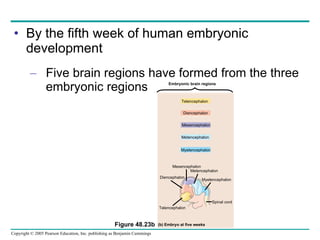
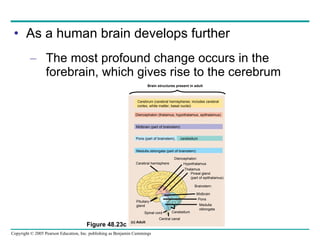
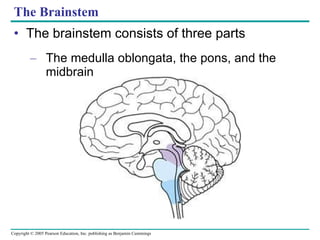
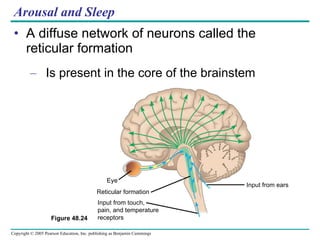
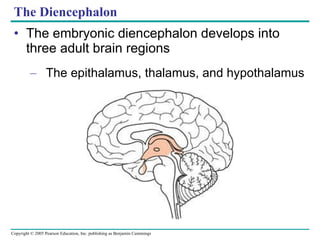
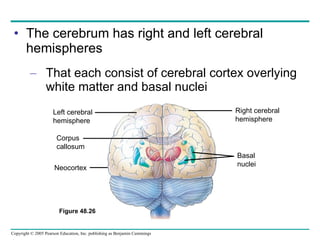
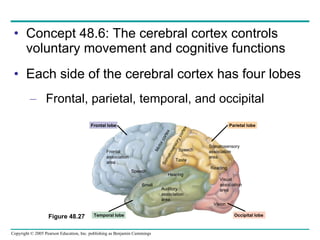
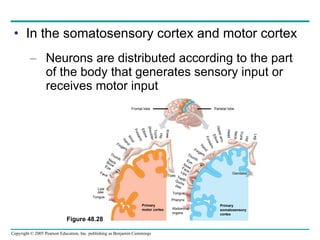
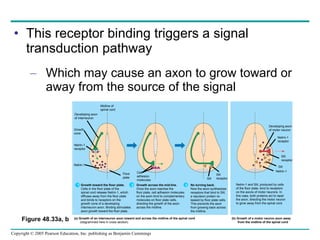
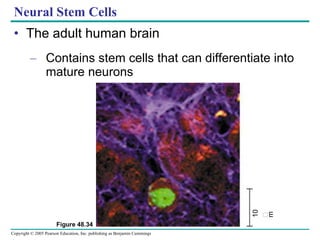
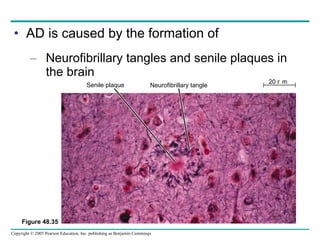
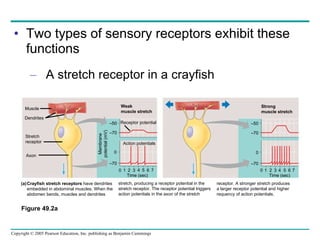
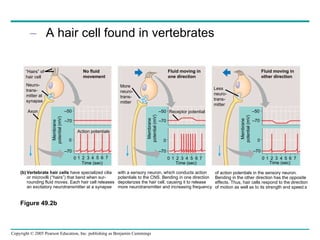
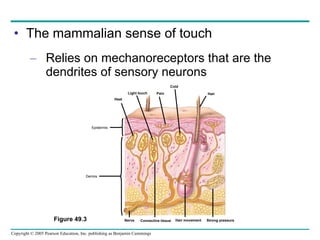
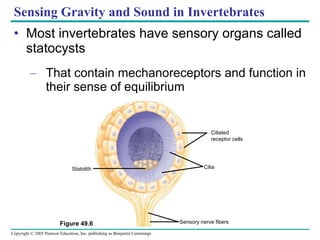
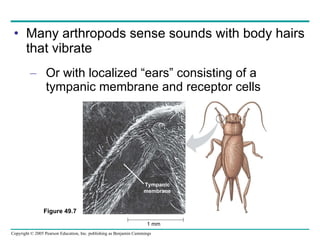
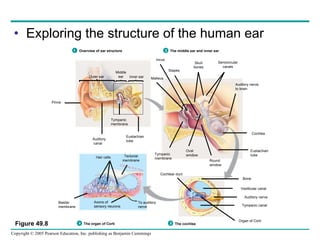
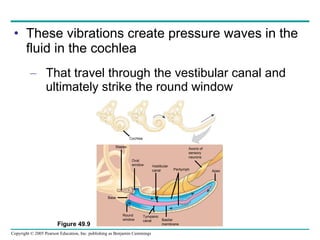
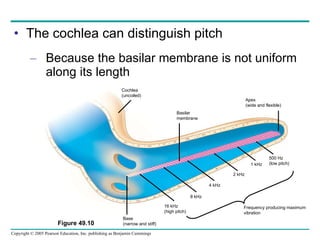
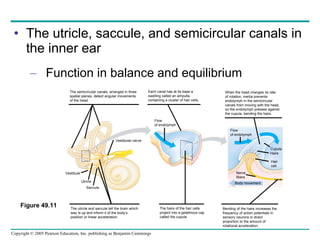
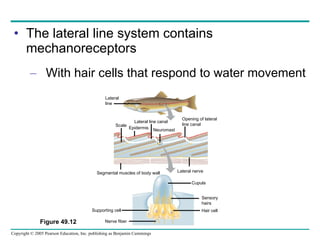
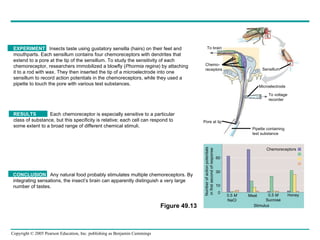
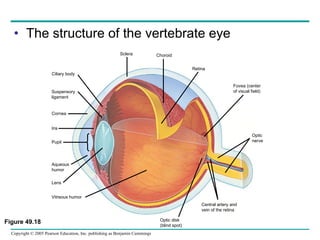
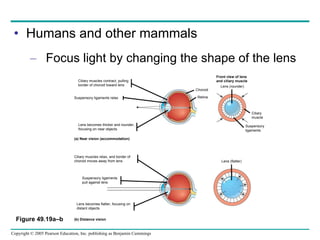
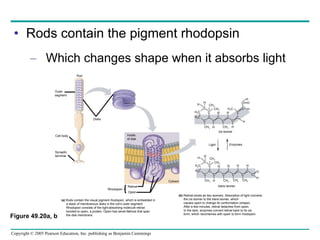
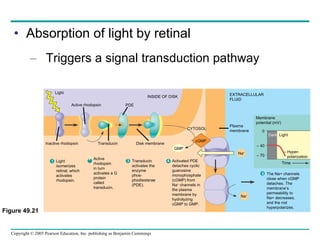
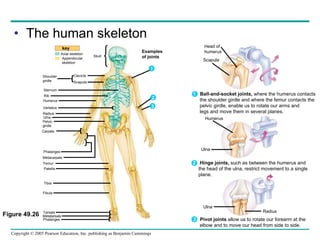
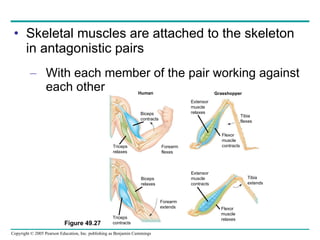
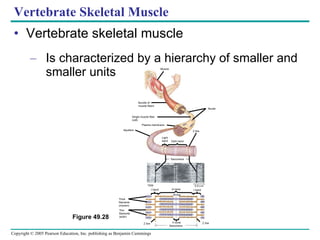
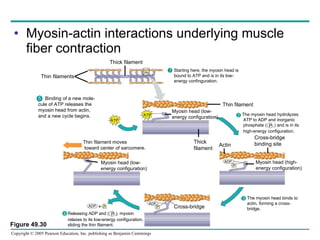
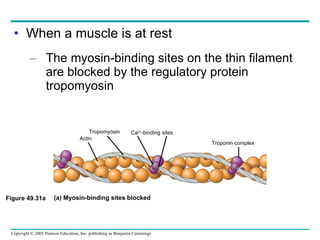
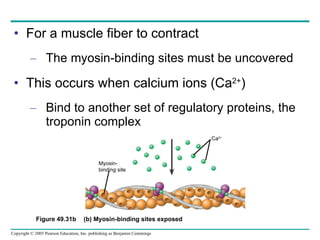
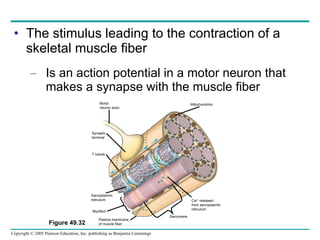
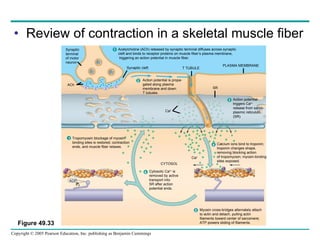
The document summarizes key concepts about nervous systems. It describes how neurons are organized into circuits in different animal groups, from nerve nets to central and peripheral nervous systems. It explains the three stages of information processing - sensory input, integration in the central nervous system, and motor output. It also describes how action potentials are generated and conducted along axons through the opening and closing of voltage-gated ion channels.










![In all neurons, the resting potential Depends on the ionic gradients that exist across the plasma membrane CYTOSOL EXTRACELLULAR FLUID [Na + ] 15 m M [K + ] 150 m M [Cl – ] 10 m M [A – ] 100 m M [Na + ] 150 m M [K + ] 5 m M [Cl – ] 120 m M – – – – – + + + + + Plasma membrane Figure 48.10](https://image.slidesharecdn.com/48-nervoustext-110121014754-phpapp02/85/48-nervous-text-24-320.jpg)



































































































































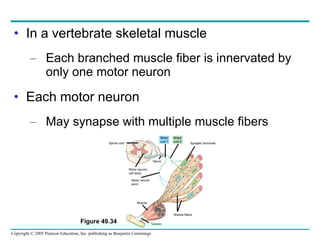




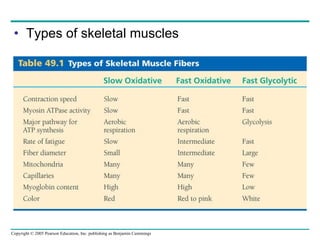







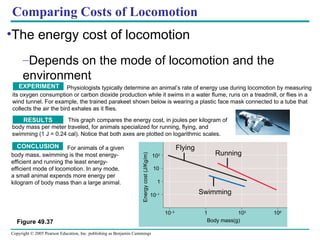



























































![[12] ANAPHYSIO The Nervous System: Nervous Tissue](https://cdn.slidesharecdn.com/ss_thumbnails/anaphysioch121-221210142254-93789f95-thumbnail.jpg?width=640&height=640&fit=bounds)

![12 [chapter 12 nervous tissue]](https://cdn.slidesharecdn.com/ss_thumbnails/12chapter12nervoustissue-170828041102-thumbnail.jpg?width=640&height=640&fit=bounds)















