Downloaded 36 times







































































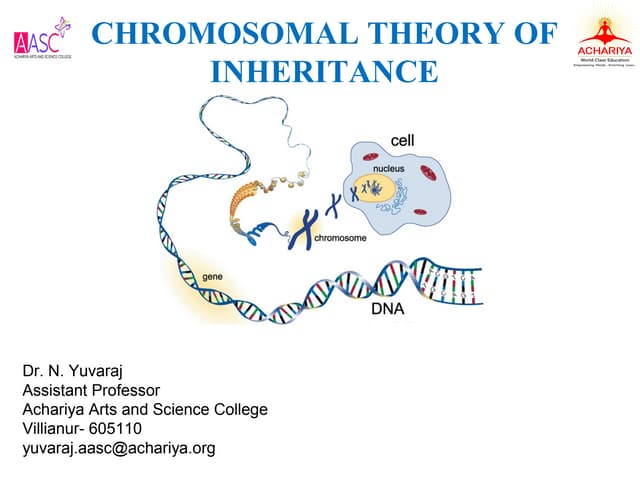
This document discusses meiosis and genetic linkage in plants and animals. It provides details on: - The alternation of generations life cycle in plants, which involves a diploid sporophyte and haploid gametophyte stage. - How crossing over during meiosis increases genetic variation by exchanging parts of homologous chromosomes. - How the frequency of recombination between two genes indicates their distance on the same chromosome, and is used to construct genetic linkage maps.