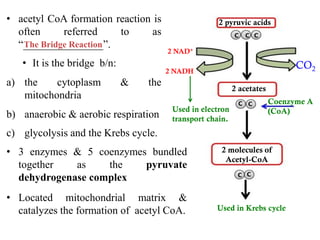
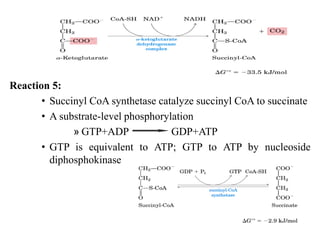
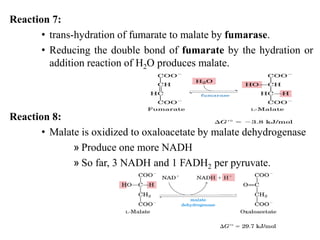
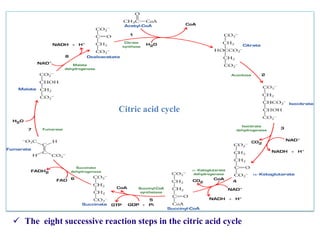
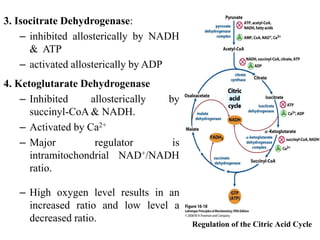
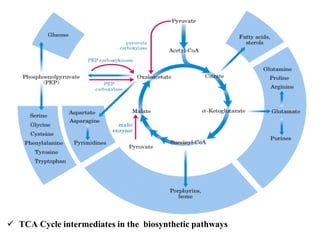
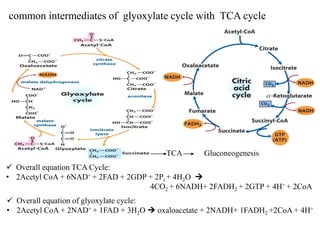
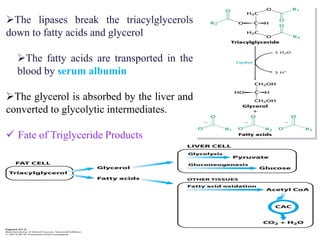
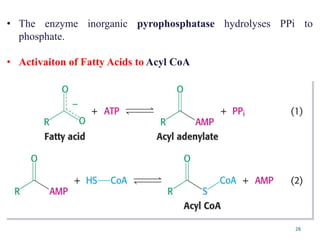
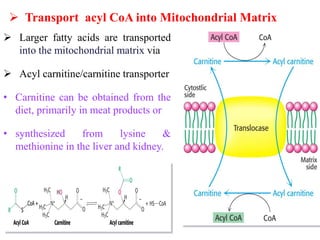
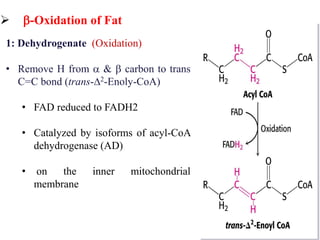
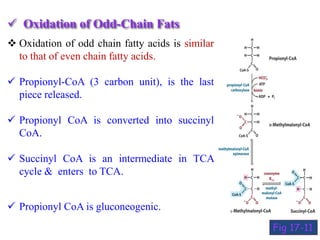
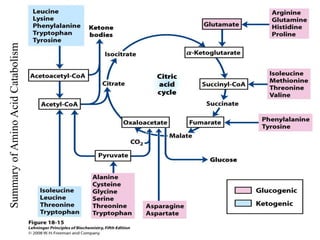
The citric acid cycle is the final stage of breaking down nutrients and occurs in the mitochondria. It uses acetyl-CoA molecules to produce carbon dioxide, hydrogen atoms, and ATP through a set of 8 enzyme-catalyzed reactions. First, pyruvate is converted to acetyl-CoA, which then feeds into the citric acid cycle to be oxidized and generate energy. The cycle is regulated by the energy levels of the cell and produces precursors for biosynthesis.
















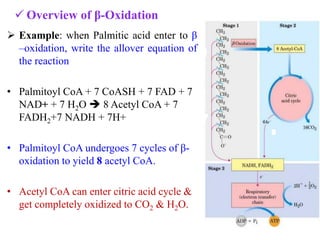
![Energetics of β- Oxidation
Mechanism ATP yield
I. β- Oxidation 7 cycles
7 FADH2 [Oxidized by Electron Transport Chain (ETC) each
FADH2 gives 2 ATP ]
7 NADH (Oxidized by ETC, each NADH
Liberate 3ATP)
14
21
II. From 8 Acetyl CoA
Oxidized by citric acid cycle, each acetyl CoA
provides 12
96
Total energy from one molecule of palmitoyl CoA
Energy utilized for activation
(Formation of palmitoyl Co A)
131
-2
Net yield of oxidation of one molecule of palmitate =129](https://image.slidesharecdn.com/thecitricacidcycle1-230516202238-10c74d1d/85/The_citric_acid_cycle-1-pdf-36-320.jpg)



















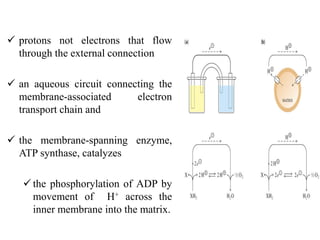








![7.9.3. Regulation of Oxidative Phosphorylation
• the mass-action ratio of the ATP-ADP system ([ATP]/([ADP][Pi]).
• some energy-requiring process (eg. Protein synthesis) increases,
the rate of breakdown of ATP to ADP and Pi increases,
• lowering the mass-action ratio.
• With more ADP available for oxidative phosphorylation, the rate
of respiration increases, causing regeneration of ATP.
An inhibitory protein (IF1) prevents ATP hydrolysis during
Ischemia.
• When a cell is ischemic (deprived of oxygen), as in a heart
attack or stroke,](https://image.slidesharecdn.com/thecitricacidcycle1-230516202238-10c74d1d/85/The_citric_acid_cycle-1-pdf-79-320.jpg)
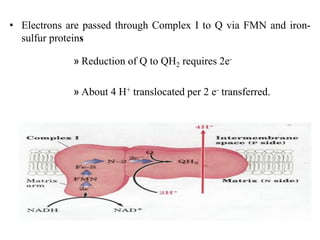
![3. Electron Transport System (ETS) Con.t
• electron transfer to oxygen ceases, and so does the pumping of
protons.
• The proton-motive force soon collapses.
• the ATP synthase operate in reverse, hydrolyzing ATP to pump
protons outward [ATP]↓
• protein inhibitor, IF1, a small (84 amino acids), binds to two ATP
synthase and inhibit their ATPase activity.
• IF1 is inhibitory only in its dimeric form, which is favored at pH
lower than 6.5.](https://image.slidesharecdn.com/thecitricacidcycle1-230516202238-10c74d1d/85/The_citric_acid_cycle-1-pdf-80-320.jpg)

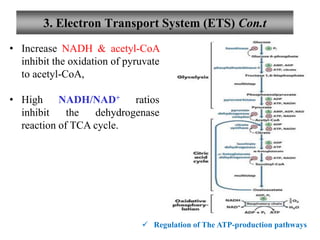
![3. Electron Transport System (ETS) Con.t
Regulation of The ATP-production pathways
• ATP-producing pathways are coordinately regulated.
• Interlocking regulation of glycolysis, pyruvated oxidation, the
citric acid cycle ,and oxidation phosphorylation by
• the relative [ ] of ATP, ADP, and AMP, & by NADH.
• All 4 pathway are accelerate when the use of ATP & the formation
of ADP, AMP, and Pi increase.
• Interlocking of glycolysis & TCA cycle by citrate, inhibits
glycolysis,
• supplements the action of the adenine nucleotide system](https://image.slidesharecdn.com/thecitricacidcycle1-230516202238-10c74d1d/85/The_citric_acid_cycle-1-pdf-82-320.jpg)