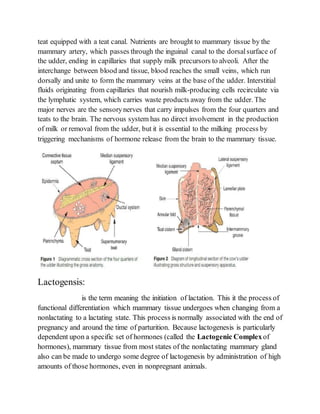
1. The document discusses milk synthesis and lactation in mammals. It describes the anatomy and physiology of the mammary gland and the processes of lactogenesis, galactopoiesis, and milk synthesis.
2. Lactogenesis involves the differentiation of mammary epithelial cells from a non-secretory to a secretory state in two stages: initial cytological changes and limited milk synthesis before birth, followed by copious milk secretion around the time of birth.
3. Milk synthesis occurs through the uptake of nutrients from blood by the mammary epithelial cells, which synthesize milk components like proteins, lactose, and fat within the cells and secrete them into the alveolar lumen for removal as milk.




![processing of milk proteins, synthesis of lactose, and the osmotic draw for
water. The Golgi apparatus is very important to the synthesis of skim milk
components. Note that lactose (and therefore much of the water of milk) is
secreted via the secretory vesicles along with the milk proteins.
Milk fat precursors to milk fat : Precursors of milk fat synthesis are also
taken up by the epithelial cells at the basolateral membrane. Acetate and ß-
hydroxybutyrate are important precursors of fatty acid synthesis in mammary
cells in some species (ruminants, especially). These precursors are absorb
through the basolateral membrane. In addition, preformed fatty acids,
glycerol, and monoacylglycerides are absorbed at the basolateral membrane.
All these components enter into the synthesis of triglycerides of milk (see
milk Fat Lesson). Milk fat triglycerides are synthesized on the smooth
endoplasmic reticulum (SER) and form small droplets. Numerous small lipid
droplets will fuse together as the growing lipid droplet moves toward the
apical membrane. At the apical membrane the large lipid droplet forces out
the apical membrane of the cell, the apical membrane surrounds the lipid
droplet until it pinches off and enters the lumen. [Imagine standing inside a
balloon and trying to punch your hand through the balloon's wall. The
balloon's wall would wrap around your hand.] So, in the lumen of the
alveolus, the milk fat globule (or milk lipid globule as it is now called) is
surrounded by a membrane. This membrane originally was part of the
epithelial cell's apical membrane. Note that INSIDE the cell the lipid is NOT
membrane bound and is called a lipid droplet, while after secretion in the
LUMEN, the milk lipid globules are surrounded by a membrane.](https://image.slidesharecdn.com/assignment-190930163730/85/MILK-SYNTHESIS-and-lactation-6-320.jpg)













































































