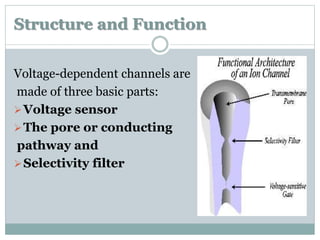
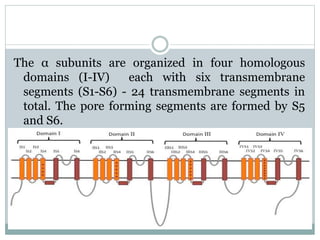
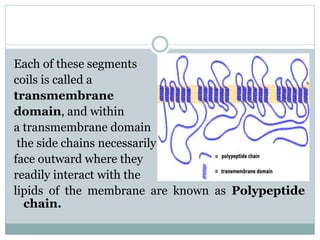
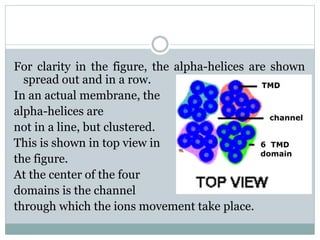
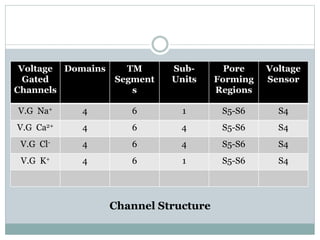
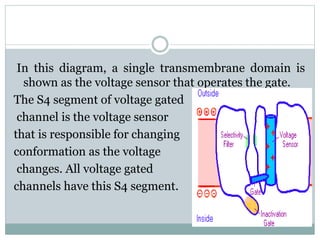
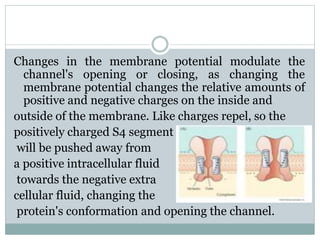
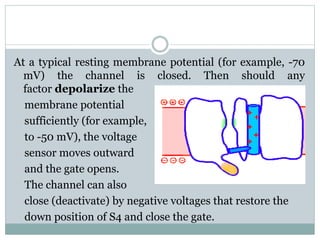
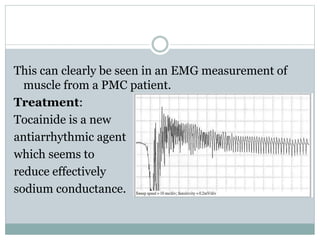
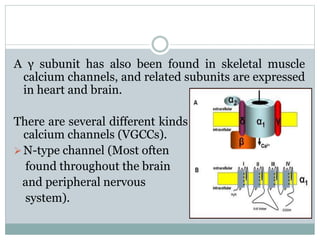
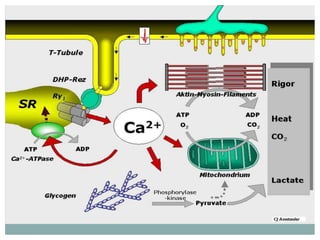
Ion channels are membrane proteins that regulate ion flow, essential for electrical signaling in cells. They can be classified into voltage-gated and ligand-gated channels, playing vital roles in various physiological functions and diseases. Recent studies have highlighted their importance in drug development, with specific channelopathies linked to genetic disorders and toxins.










































![ http://genetics.thetech.org/original.
Dinarello CA, Porat R. Fever and hyperthermia. In: Fauci A,
Kasper D, Longo DL, et al, eds. Harrison's Principles of
Internal Medicine. 17th ed. [online version]. New York, NY:
McGraw Hill;2008:chap 17.
Felix Luessi, MD; Volker Siffrin; Frauke Zipp.
Neurodegeneration in Multiple Sclerosis: Novel Treatment
Strategies: Therapeutic Approaches to Neuronal
Degeneration in MS. Medscape. 2013.
Yu-Qing Cao. Voltage-gated calcium channels and pain.
Pain. 126 (2006) 5–9.](https://image.slidesharecdn.com/ionchannels-141024171944-conversion-gate02/85/Ion-channels-types-and-their-importace-in-managment-of-diseases-56-320.jpg)
























































































