IMUNOLOGIJAIMUNOLOGIJA
IMUNITET –imunitas (zaštitasenatora u starom Rimu od sudskih
procesa tokom mandata)
-u užem smislu: otpornost prema infektivnim bolestima
-u širem smislu: zaštita organizma od infektivnih i neinfektivnih
agenasa- A N T I G E N I
IMUNSKI SISTEM – skup organa, tkiva, ćelija i njihovih produkata
koji učestvuju u imunitetu
IMUNSKI ODGOVOR – koordinisano dejstvo komponenti imunskog
sistema u borbi protiv infektivnih mikroorganizama i ostalih
štetnih agenasa
IMUNOLOGIJA – nauka koja proučava imunski sistem i imunski
odgovor
www.belimantil.info
2.
Pojam imunitet sepominje u 5.veku p.n.e. u grčkim zapisima
Znatno ranije se koncept imuniteta pominje u kineskim
zapisima: inhalacija praha dobijenog iz krusti obolelih od
velikih boginja
Edvard Džener (1798.god): sadržaj pustule inficirane virusom
kravljih boginja dat u cilju prevencije infekcije virusom velikih
boginja- pojam vakcinacije (vacca-krava; vaccinus- od krave)
ODBRANA OD INFEKTIVNIH MIKROORGANIZAMA
-imunodeficijencije, AIDS
TRANSPLANTACIJSKA IMUNOLOGIJA
TUMORSKA IMUNOLOGIJA
REAKCIJE PREOSETLJIVOSTI (ALERGIJE)
AUTOIMUNSKE REAKCIJE
www.belimantil.info
3.
ZNAZNAČČAJ IMUNOLOGIJE ZAFARMACIJUAJ IMUNOLOGIJE ZA FARMACIJU
• Terapijska primena lekova koji utiču na imunski odgovor (mehanizmi
dejstva imunosupresiva i imunopotencijatora)
• Razumevanje mehanizama preosetljivosti na lekove
• Kreiranje vakcina i razumevanje mehanizama njihovog delovanja
• Primena imunoloških metoda u dijagnostici (imunohemijske i druge)
• Kreiranje novih imunomodulatornih lekova i vakcina
• Produkcija rekombinantnih proteina za potrebe dijagnostike i terapije
(primer rekombinantni citokini)
• Produkcija poliklonskih i monoklonskih antitela za potrebe dijagnostike i
terapije
FARMACEUTSKA (IMUNO)BIOTEHNOLOGIJA
proizvodi za dijagnostiku i terapiju
www.belimantil.info
4.
TIPOVI IMUNITETATIPOVI IMUNITETA
Učestvujeu ranoj fazi odbrane
od infekcija (aktivan u prvih
nekoliko časova)
• Uvek prisustan u zdravih osoba
• Sprečava prodor mikroorganizama
u organizam i brzo eliminiše
mikrorganizme
•Stimulisan mikroorganizmima koji su
prodrli u tkivo
•Adaptira se na prisustvo
mikroorganizama
•Razvija se sporije, i
•Posreduje u kasnijoj ali efikasnijoj
fazi odbrane (razvija se posle
nekoliko dana)
Stečeni imunitet
(adaptivni, specifični)
Urođeni imunitet
(prirodni)
www.belimantil.info
5.
Komponente urođenog imuniteta
•Epitelne barijere i solubilni produkti epitela (prirodni antibiotici)
• Fagociti i prirodne ćelije ubice (NK ćelije), i njihovi produkti-citokini
• Proteini plazme (sistem komplementa)
www.belimantil.info
6.
Karakteristike urođenog imuniteta
•Urođeni imunitet prepoznaje i reaguje na mikroorganizme, ali ne i na
neinfektivne strane supstance i ćelije
• U prepoznavanju se koriste isti mehanizmi za različite vrste mikroorganizama
• Urođeni imunitet pospešuje delovanje stečenog imuniteta
www.belimantil.info
7.
Komponente stečenog imuniteta
•Limfociti i njihovi produkti:
B limfociti nosioci humoralnog imuniteta
T limfociti nosioci ćelijskog imuniteta)
• Stečeni imunitet prepoznaje mikrobske i nemikrobske molekule koji se
zajedničkim imenom nazivaju antigeni
• Prepoznavanje antigena je specifično: receptori za antigen na limfocitima
www.belimantil.info
8.
STALNA KOOPERACIJA IZMEĐUUROĐENOG I
STEČENOG IMUNITETA ČINI OSNOVU USPEŠNOG
IMUNSKOG ODGOVORA
www.belimantil.info
9.
KARAKTERISTIKE STEKARAKTERISTIKE STEČČENOGIMUNITETAENOG IMUNITETA
S P E C I F I Č N O S T
• Sposobnost specifičnog odgovora u odnosu na različite antigene
(anitgenske determinante)- specifični receptori za antigen na limfocitima
R A Z N O L I K O S T
• Sposobnost imunog sistema da odgovori na veoma veliki broj različitih
antigena- antigenski repertoar (skup svih limfocita/klonova različitih
specifičnosti)
• K l o n l i m f o c i t a
Limfociti identične specifičnosti za neki antigen (antigensku
determinantu) koji potiču od iste ćelije
www.belimantil.info
10.
KARAKTERISTIKE STEKARAKTERISTIKE STEČČENOGIMUNITETAENOG IMUNITETA
P A M Ć E N J E
• Stvaranje dugoživećih limfocita pamćenja nakon primarnog
imunskog odgovora
• Snažniji imunski odgovor na ponovnu ili perzistentnu infekciju
S P E C I J A L I Z A C I J A
• Optimalni imunski odgovor prema različitim vrstama
mikroorganizama/antigena (primer ćelijski i humoralni)
NEREAKTIVNOST NA SOPSTVENE ANTIGENE-
autotolerancija
SAMOOGRANIČAVANJE
prevencija oštećenuja tkiva tokom reakcije na antigene iz
spoljašnje sredine
www.belimantil.info
Memorijski limfociti
PRIMARNI ISEKUNDARNI IMUNI ODGOVOR
Memorijski limfocitiEfektorski limfociti
Klonovi limfocita
Efektorski limfociti
antigen
antigen
PRIMARNI IMUNI ODGOVOR
SEKUNDARNI IMUNI ODGOVOR
www.belimantil.info
14.
HUMORALNI IMUNITET
posredovan antitelimakoja su produkti B
limfocita
učestvuje u zaštiti od ekstracelularnih
mikroorganizama neutralizacijom
mikroorganizama i njihovih tokisina
www.belimantil.info
15.
ĆELIJSKI IMUNITET
posredovan Tlimfocitima
učestvuje u odbrani od intraćelularnih
mikroorganizama
Aktivacijom fagocita
Aktivacijom citotoksičnih limfocita
www.belimantil.info
16.
AKTIVNI IMUNITET
– indukovaninfekcijom ili vakcinacijom
PASIVNI IMUNITET
– prenet antitelima ili limfocitima iz osobe
sa aktivnim imunitetom u neimunizovanu
osobu (imunitet novorođenčeta prenet od
majke preko placente)
www.belimantil.info
• Učestvuje uranoj fazi odbrane od infekcija (aktivan u prvih
nekoliko časova)
• Uvek prisustan u zdravih osoba
• Sprečava prodor mikroorganizama u organizam i brzo eliminiše
mikrorganizme koji su prodrli u tkivo
Komponente urođenog imuniteta
• Epitelne barijere
• Fagociti i prirodne ćelije ubice (NK ćelije)
• Proteini plazme (sistem komplementa, C reaktivni
protein, citokini prirodnog imuniteta)
UROUROĐENIĐENI IMUNITETIMUNITET
www.belimantil.info
20.
• Urođeni imunitetprepoznaje i reaguje na mikroorganizme ali
ne i na neinfektivne strane supstance
• Urođeni imunitet ne reaguje prema sopstvenim ćelijama zbog
postojanja regulatornih molekula
• Komponente urođenog imuniteta koriste iste mehanizme u
prepoznavanju različitih vrsta mikroorganizama
• Odgovor urođenog imuniteta je isti pri ponovnom kontaktu sa
mikrobima
• Urođeni imunitet pospešuje delovanje stečenog imuniteta
Karakteristike urođenog imunitetaKarakteristike urođenog imuniteta
www.belimantil.info
21.
Prepoznavanje
struktura koje su
zajedničkeza više
vrsta mikroorganizama
a nisu prisutne na
vlastitim ćelijama
(LPS; manoza na glikoproteinima;
dvolančana RNK virusa;
nemetilovani CpG nukleotidi)
Shema preuzeta iz knjige”Basic Immunology”, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
22.
Prepoznavanje struktura
koje suvažne za
preživljavanje
mikroorganizama
Receptori koji prepoznaju
strukture mikroorganizama
su kodirani genima matične
loze koji ne podležu
procesima rekombinacije
Shema preuzeta iz knjige”Basic Immunology”,
Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
23.
Primeri koji ilustrujusposobnost urođenog imuniteta da
reguje sa mikroorganizmima ali ne i sa sopstvenim
ćelijama
Molekuli koje prepoznaju ćelije urođenog imuniteta nisu
prisutni na sopstvenim ćelijama
NK ćelije ne vrše citolizu sopstvenih ćelija zbog ispoljavanja
inhibitornih receptora
Komplement ne oštećuje sopstvene ćelije zbog protektivnog
delovanja regulatornih proteina komplementa
www.belimantil.info
24.
Fizička zaštita
Produkcija peptidnihantibiotika
(defenzini)
γδ+ T limfociti (intraepitelni limfociti)
(prepoznavanje lipida mikroorganizama)
B1 limfociti (peritoneum)
(produkcija prirodnih antitela IgM klase)
Shema preuzeta iz knjige”Basic Immunology”, Abbas i Lichtman, isključivo
za potrebe predavanja
www.belimantil.info
25.
- Ćelije mijeloidneloze (granulociti)
- Prve ćelije koje odgovaraju na infekciju
bakterijama i gljivicama
- Fagocitoza u cirkulaciji
- Ekstravazacija u tkivo
- Kratkoživeće ćelije
NEUTROFILI
www.belimantil.info
26.
MONOCITI
subpopulacije makrofaga u
limfoidnimorganima
histiociti
osteoklastiKupferove ćelije mikroglia alveolarne
makrofage
AKTIVACIJA
džinovske ćelije epiteloidne ćelije
-Zajedničko poreklo sa granulocitima
-Diferentovanje od monoblasta u kostnoj srži
-Migracija u tkivo i transformacija u
makrofage
www.belimantil.info
PREPOZNAVANJEPREPOZNAVANJE
RECEPTORI:RECEPTORI:
- manozni receptor
-N-formil metionil receptor
- Receptor “čitač”
- Mac1 (CD11b/CD18)
- Toll-slični receptori
- Receptori za Fc fragment antitela
- Receptori za komponente
komplementa
FAGOCITOZA I FORMIRANJE
FAGOZOMA
- Fuzija fagozoma i lizozoma (formiranje
fagolizozoma)
- Aktivacija enzima: fagocitna oksidaza
(superoksid anjon; slobodni radikali),
NO- sintaza (NO), lizozomalne
proteaze
UBIJANJE MIKROORGANIZMA
Oštećenje tkiva zbog oslobađanja enzima
u tkivo
(pospešavanje inflamacije)
Shema preuzeta iz knjige”Basic Immunology”, Abbas i Lichtman,
isključivo za potrebe predavanja
www.belimantil.info
29.
Shema preuzeta izknjige”Basic Immunology”, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
30.
Stimulacija endotela Prodormikroorganizma u tkivo
ekspresija adhezivnih molekula - fagocitoza od strane tkivnih makrofaga
* P selektin - aktivacija tkivnih makrofaga
* E selektin (produkcija TNFa, IL-1, hemokina)
produkcija hemokina
Shema preuzeta iz knjige”Basic Immunology”, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
31.
Vezivanje leukocita zaadhezivne molekule putem liganada za selektine (slaba veza - “kotrljanje” leukocita)
Aktivacija Mac1 (CD11b/CD18) integrina
Vezivanje za integrinske ligande na endotelu (stabilna veza)
Transmigracija u tkivo (pod uticajem hemokina)
INFLAMACIJA (ZAPALJENJE)
nakupljanje leukocita u tkivu
povećana propustljivost i proširenje krvnih sudova
www.belimantil.info
32.
TNFα Aktivacija endotelai neutrofila, inflamacija
povećanje telesne temperature, sinteza proteina
akutne faze u jetri, povećanje katabolizma u
mišićima i masnom tkivu, indukcija apoptoze
brojnih ćelija.
IL-1 Aktivacija endotela i neutrofila, inflamacija
povećanje telesne temperature, sinteza proteina
akutne faze u jetri
HEMOKINI CCL i CXCL familija
aktivacija i hemotaksa leukocita
Shema preuzeta iz knjige”Basic Immunology”, Abbas i Lichtman, isključivo
za potrebe predavanjawww.belimantil.info
33.
IL-6 Proliferacija efektorskihB limfocita,
sinteza proteina akutne faze u jetri, inflamacija
IL-12 Stimulacija produkcije IFN-γ od strane NK ćelija i
T limfocita, povećanje citotoksične aktivnosti T
limfocita, diferencijacija Th1 ćelija
IL-15 Proliferacija NK ćelija i T limfocita
IL-18 Stimulacija produkcije IFN-γ
www.belimantil.info
34.
Interferoni (tip I)
IFN-α,IFN-β Anti virusna aktivnost, povećanje
ekspresije MHC molekula I klase,
aktivacija NK ćelija
Interferon (tip II)
IFN-γ Aktivacija makrofaga, stimulacija
produkcije pojedinih potklasa IgG
www.belimantil.info
35.
- Veliki granuliranilimfociti (LGL)
- Nemaju receptor za antigen
- Čine oko 10% svih limfocita u krvi
- Učestvuju u nespecifičnoj citotoksičnosti
- ubijaju zaražene ćelije
- tumorske ćelije
- alogene ćelije
- sopstvene izmenjene ćelije
NK ćelije
Shema preuzeta iz knjige”Basic Immunology”, Abbas i Lichtman, isključivo za
potrebe predavanja
-Produkuju IFN-γ nakon stimulacije od strane IL12
koga produkuju makrofagi
-Vezuju antitela i učestvuju u citotoksičnosti
zavisnoj od antitela (ADCC)
-Prepoznaju ciljne ćelije posle deaktivacije
inhibitornih receptora
-Citotoksičnost zavisna od direktnog kontakta
- Slični citolitički mehanizmi kao i kod CTL
www.belimantil.info
36.
Shema preuzeta izknjige”Basic Immunology”,
Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
37.
PODSETNIK
1. Imunologija, imunitet,imuni sistem, imuni odgovor
2. Imuni odgovor:
- odbrana od infektivnih agenasa
(mikroorganizama)
- odbrana od neinfektivnih agenasa (stranih
supstanci, antigena)
- reakcije prema stranim ćelijama i tkivima
(imunologija transplantacije)
- reakcije prema tumorskim ćelijama
(tumorska imunologija)
3. Urođeni (prirodni, nespecifični) imunitet:
rana faza odbrane organizma, uvek prisutan kod svih
osoba
www.belimantil.info
38.
4. Stečeni (adaptivni,specifični) imunitet:
- kasnija faza odbrane; specifičan u odnosu na antigen,
specijalizovane vrste reakcija koje na najefikasniji
način uklanjaju antigen;
5. Zajedničko delovanje urođenog i stečenog imuniteta
6. Komponente urođenog imuniteta:
- epitelne barijere (koža i sluznice)
- fagociti, NK ćelije (prirodne ubice)
- sistem komplementa
- citokini
7. Komponente stečenog imuniteta:
- limfociti B i limfociti T
- solubilni produkti (antitela, citokini)
www.belimantil.info
39.
8. Tipovi stečenogimuniteta:
- humoralni (B limfociti sekretuju antitela): odbrana
od ekstraćelijskih mikroorganizama;
- celularni (T limfociti i njihovi produkti): odbrana
od intraćelijskih mikroorganizama;
9. Vrste stečenog imuniteta:
- aktivni (prirodni, veštački)
- pasivni (prirodni, veštački)
10. Primarni i sekundarni specifični imuni odgovor
11. Faze specifičnog imunog odgovora: prepoznavanje,
aktivacija, efektorska faza, zaustavljanje, pamćenje
www.belimantil.info
1. L IM F O C I T I
Ključne ćelije specifičnog imuniteta
Jedine ćelije sa receptorom za antigen
B limfociti (humoralni imunitet)
T limfociti (ćelijski imunitet)
-CD4 limfociti (T limfociti pomoćnici)
-CD8 limfociti (T citotoksični limfociti)
www.belimantil.info
43.
SLISLIČČNOSTI I RAZLIKEIZMEĐUNOSTI I RAZLIKE IZMEĐU
T i B LIMFOCITAT i B LIMFOCITA
S L I Č N O S T I
Slična morfologija
Slična struktura receptora za antigen i njihovih gena
Stvaranje limfocita pamćenja
Recirkulacija
www.belimantil.info
44.
R A ZL I K E
Različito mesto stvaranja
B limfociti . . . . . . kostna srž
T limfociti . . . . . . timus
Različito ispoljavanje (ekspresija) membranskih proteina
(CD molekula)
B limfociti...............BCR, CD19, CD20, CD21
T limfociti...............TCR, CD3, CD4, CD8
www.belimantil.info
45.
Različit oblik ipriroda antigena koje vezuju posredstvom receptora
za antigene
B limfociti vezuju: nativne, solubilne antigene
antigene ćelija (nesolubilne)
proteinske i neproteinske antigene
T limfociti vezuju: samo proteinske antigene prerađene do peptida i ispoljene
u sklopu jednog drugog molekula (MHC) na membrani
specijalizovanih ćelija (APC)
Različite efektorske funkcije
B limfociti: - produkcija antitela
T limfociti: - produkcija citokina (T limfociti pomoćnici, Th-helper)
- citoliza ciljnih ćelija (citotoksični T limfociti, CTL)
R A Z L I K E
www.belimantil.info
46.
Shema preuzeta izknjige Basic Immunlogy, Abbas i Lichtman, isključivo za potrebe predavanjawww.belimantil.info
47.
2. ANTIGEN2. ANTIGEN--PREZENTUJUPREZENTUJUĆĆEEĆĆELIJE (APC)ELIJE (APC)
Definicija:
- Ćelije koje imaju sposobnost hvatanja, prerade i prikazivanja
(prezentacije) antigena u odgovarajućoj formi limfocitima.
- Ako obezbeđuju drugi signal za aktivaciju limfocita
(kostimulatorni signal) onda su to profesionalne APC
- Najznačajnije profesionalne APC
DENDRITIČNE (DENDRITSKE) ĆELIJE
www.belimantil.info
48.
DENDRITIČNE ĆELIJE
Dendritična morfologija
Hvatanje,prerada i prezentacija antigena
Zrele i nezrele
Mijeloidne i plazmocitoidne
Slike preuzete iz knjige Basic Immunology, Abbas i Lichtman
isključivo za potrebe predavanja
www.belimantil.info
49.
NEZRELE DC :NEZRELEDC : hvatanje i prerada antigenahvatanje i prerada antigena
KRV
LIMFA
EPIDERMIS KOŽE (LANGERHANSOVE ĆELIJE) I EPITEL
SLUZNICA
PERIFERNA TKIVA (INTRESTICIJUM)
ZRELE DCZRELE DC :: prezentacija antigena T limfocitima iprezentacija antigena T limfocitima i
kostimulacijakostimulacija
L. ČVOR (PARAKORTKS)
SLEZINA (PALS)
FOLIKULARNE ( FOLIKULSKE) DENDRITIFOLIKULARNE ( FOLIKULSKE) DENDRITIČČNENE ĆĆELIJEELIJE
FOLIKULI SEZINE I LIMFNIH ČVOROVA
PREZENTACIJA ANTIGENA B LIMFOCITIMA
ZNAČAJ ZA AFINITETNO SAZREVANJE B LIMFOCITA
www.belimantil.info
50.
3. EFEKTORSKE3. EFEKTORSKEĆĆELIJEELIJE
B LIMFOCITI (PLAZMOCITI): Produkcija antitela
T LIMFOCITI: CD4+ T (produkcija citokina)
CD8+ T (specifična citotoksičnost
NK ĆELIJE: nespecifična citotoksičnost
MAKROFAGI: urođeni imunitet
efektorske ćelije specifičnog imuniteta
NEUTROFILI: urođeni imunitet
efektorske ćelije specifičnog imuniteta
www.belimantil.info
51.
ORGANI I TKIVAIMUNOG SISTEMAORGANI I TKIVA IMUNOG SISTEMA
ORGANI: - CENTRALNI (primarni ili generativni):
(nastanak i razvoj limfocita)
timus
kostna srž
- PERIFERNI (sekundarni)
(aktivacija zrelih limfocita)
slezina
limfni čvorovi
TKIVA: ORGANIZOVANO LIMFOIDNO TKIVO
- digestivnog trakta (GALT)
- krajnici (tonzile), Pajerove ploče, apendiks
- bronhopulmonalnog trakta (BALT)
DIFUZNO LIMFOIDNO TKIVO
- koža, sluznice
www.belimantil.info
52.
HEMATOPOEZA UHEMATOPOEZA UKOSTNOJ SRKOSTNOJ SRŽŽII
Kostna srž je mesto hematopoeze gde nastaju sve ćelije krvi. Za sazrevanje ovih ćelija,
uključujući i limfocite, bitni su faktori mikrosredine (ćelije strome i solubilni faktori rasta i
diferencijacije)
Shema preuzeta iz knjoge Basic Immunology, Abbas i Lichtman isključivo za potrebe predavanjawww.belimantil.info
53.
TIMUSTIMUS
1. Centralni limfoidniorgan za razvoj T
limfocita
2. Sastavljen iz dva režnja koji su vezivnim
pregradama podeljeni na režnjiće
3. Svaki režnjić se sastoji iz spoljašnjeg
(tamnijeg) dela- korteks ili kora i
unutrašnjeg (svetlijeg) dela- srž ili medula
4. Ćelije timusa su limfoidne (timociti) i
nelimfoidne (ćelije mikrosredine); timociti su
timusni T limfociti različitog stepena zrelosti
5. Ćelije mikrosredine su epitelne ćelije,
makrofagi, dendritične ćelije i dr. Ove ćelije i
njihovi produkti obezbeđuju optimalne
uslove za sazrevanje T limfocita i procese
selekcije
Slike preuzete iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
54.
TIMUSTIMUS
7. Epitelne ćelijeucestvuju u sazrevanju T
limfocita i procesima pozitivne i negativne
selekcije putem direktnog kontakta ili
posredstvom solubilnih faktora (timusni
hormoni, citokini)
8. Timusni makrofagi uklanjaju neselekcionisane
timocite
9. Dendritične ćelije medule učestvuju u
negativnoj selekciji timocita
Slike preuzete iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
55.
S L EZ I N AS L E Z I N A
STROMA: kapsula, krvni sudovi
PARENHIM:
- BELA PULPA
- MARGINALNA ZONA
- CRVENA PULPA
B e l a p u l p a:
1. Folikuli (B zone):
- primarni
- sekundarni (germinativni centri)
- folikularne dendritične ćelije
2. PALS- periarteriolarni limfocitni
omotač (T zone):
- T limfociti
- dendritične ćelije
Slike preuzete iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanjawww.belimantil.info
56.
M a rg i n a l n a z o n a
- T i B limfociti
- makrofagi
C r v e n a p u l p a
- venski sinusoidi
- T i B limfociti
- makrofagi
- ostale ćelije
SLEZINASLEZINA
www.belimantil.info
57.
LIMFNILIMFNI ČČVORVOR
STROMA:
kapsula, krvnisudovi, aferentni, eferentni i
intranodalni limfatički sudovi
PARENHIM:
1. K o r t e k s:
- folikuli primarni i sekundarni (B zone)
2. Duboki korteks, parakorteks, parafolikularni
korteks (T zone):
- T limfociti
- dendritične ćelije
- venule sa visokim endotelom (HEV)
3. M e d u l a
Slike preuzete iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanja
www.belimantil.info
58.
Limfociti ulaze ulimfni čvor preko
postkapilarnih venula sa visokim endotelom
Migracija T i B limfocita u odgovarajuće
regione je pod uticajem specifičnih hemokina
koje produkuju nelimfoidne ćelije tih regiona
Dendritične ćelije koje su prihvatile antigen u
epitelu, migriraju preko aferentnih limfatičnih
sudova u T zone limfnog čvora
Slike preuzete iz knjige Basic Immunology, Abbas i Lichtman,
isključivo za potrebe predavanja
www.belimantil.info
59.
Naivni T limfocitimigriraju iz krvi preko postkapilarnih venula sa visokim endotelom u
predeo parakorteksa, gde prepoznaju antigen ispoljen na membrani dendritične ćelije;
Aktivirani T limfociti napuštaju limfni čvor preko eferentnih limfatičnih sudova i preko
ductus thoracicus ulaze u krv
Iz krvi, aktivirani T limfociti migriraju preko endotela krvnih sudova u periferna tkiva
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
60.
PODSETNIK:
1. Ćelije imunogsistema su: limfociti, antigen prezentujuće ćelije i
efektorske ćelije
2. Limfociti mogu biti: limfociti B (poreklo kostna srž), limfociti T (poreklo
timus) i NK (natural killer) ćelije (poreklo kostna srž); podvrste T
limfocita su CD4+T limfociti i CD8+T limfociti
3. Naivni limfociti i memorijski limfociti (limfociti pamćenja)
4. Ćelije koje prikazuju antigen T limfocitima u odgovarajućoj formi i
obezbeđuju drugi signal aktivacije su antigen-prezentujuće ćelije (APĆ);
najznačajnije APĆ su dendritične ćelije
5. Efektorske ćelije imunog sistema su plazmociti, T limfociti pomoćnici (T
helper ili Th), citolitični T limfociti (CTL), NK ćelije, makrofagi, neutrofili
6. Centralni(primarni, generativni) limfoidni organi su kostna srž i timus
7. Periferni (sekundarni) limfoidni organi su slezina i limfni čvorovi
8. Limfoidna tkiva su GALT i BALT, kao i difuzno tkivo kože i sluznica
9. U centralnim limfoidnim organima sazrevaju limfociti, a u perifernim
organima i tkivima zreli limfociti prepoznaju antigen i aktiviraju se
www.belimantil.info
Komponente
•LIMFOCITI
•SOLUBILNI PROIZVODI LIMFOCITA
-Antitela
- Citokini
Specifično PREPOZNAVANJE Ag
pomoću RECEPTORA ZA Ag (TCR,
BcR) na limfocitima
Različiti mehanizmi za najefikasnije
uklanjanje antigena
www.belimantil.info
B LIMFOCITI PREPOZNAJU:
•proteine
• polisaharide
• lipide
• male molekule (hapteni)
Nesolubilne antigene koji su sastavni deo
membrane ćelija, kapsule i omotača
mikroorganizama
Solubilne antigene (npr. toksini bakterija)
Nativne forme antigena
www.belimantil.info
65.
T LIMFOCITI PREPOZNAJU:
•Proteinrazgrađen do peptida, povezan
u kompleks sa MHC molekulima i
ispoljen na membrani APC
MHC restrikcija
Svaki T limfocit poseduje dvojnu specifičnost TcR:
- prema određenim reziduama peptidnog Ag
- prema određenim reziduama MHC molekula koji je
“prihvatio” taj peptidni Ag
www.belimantil.info
66.
Shema preuzet izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
67.
ANTIGENI
SPECIFIČNI IMUNI ODGOVORNA PROTEINSKE
ANTIGENE ZAPOČINJE INTERAKCIJOM TRI
MOLEKULA PRISUTNA NA MEMBRANAMA T
LIMFOCITA I ANTIGEN-PREZENTUJUĆE ĆELIJE
1. TcR (na T limfocitu)
2. Antigeni peptid/MHC molekul (na APĆ)
www.belimantil.info
ŠTA SE DEŠAVASA MIKROBNIM,
ODNOSNO PROTEINSKIM
ANTIGENOM POSLE ULASKA U
ORGANIZAM ?
•Ko prihvata antigen ?
•Kako se transportuje antigen ?
•Gde se antigen koncentriše ?
www.belimantil.info
70.
PRIMER
- Mikroorganizam prodirepreko
epitela; u tkivu ga prihvataju
dendritične ćelije (APC) koje se
normalno nalaze u tkivima.
- Dendritična ćelija prihvata antigen
putem fagocitoze (ako je antigen
ćelija-mikroorganizam) ili pinocitoze
(ako je antigen slobodan, solubilan).
- Dendritična ćelija “natovarena”
antigenom napušta epitel i putem
limfotoka odlazi do regionalnog
limfnog čvora gde se odvija imuni
odgovor.
- Solubilne antigene koji se nađu u
limfotoku prihvataju dendritične
ćelije limfnih čvorova, dok antigene
iz krvotoka prihvataju dendritične
ćelije slezine.
- Svrha opisanog prihvatanja i
transportovanja antigena od mesta
ulaska do perifernog limfnoog tkiva
je da se antigen “predstavi” T
limfocitima i time započne imuni
odgovor
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanja
www.belimantil.info
71.
•Dendritične ćelije u
tkivima(koži, sluznicama,
vezivnom tkivu) su nezrele.
One mogu da prihvate
antigen, ali nisu u stanju
da aktiviraju T limfocite.
•Dendritične ćelije u
perifernim limfnim tkivima
(limfnim čvorovima,
slezini) su funkcionalno
zrele, jer osim što
“predstavljaju” antigen T
limfocitima, one i
obezbeđuju dodatne
signale za aktivaciju T
limfocita.
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, isključivo za potrebe predavanja
www.belimantil.info
72.
- Nezrela dendritičnaćelija (DC) hvata
antigen i pod dejstvom pro-inflamacionih
citokina menja oblik (postaje obla), gubi
adhezivnost prema epitelu i ispoljava
receptore za hemotaktične faktore
(hemokine).
-Ove promene omogućavaju da DC sa
svojim antigenskim tovarom napusti
epitel i preko aferentnih limfnih sudova
usmeri svoje kretanje ka regionalnom
limfnom čvoru.
-Usmeravanje kretanja DC vrše hemokini
koje proizvode T zone limfnog čvora
-Tokom usmerenog kretanja (hemotakse),
DC sazrevaju:
ispoljavaju veće količine MHC molekula
ispoljavaju kostimulacione molekule
-U limfnom čvoru, DC prikazuju Ag T
limfocitima koji su specifični za taj
antigen.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanjawww.belimantil.info
73.
T limfociti neprestanorecirkulišu
kroz limfni čvor
APC koje nose antigen i “naivni” T
limfociti specifični za taj antigen
sreću se u limfnom čvoru
Ako su mikrobni antigeni prodrli u
organizam na bilo kom mestu,
odgovor T limfocita u limfnom čvoru
koji drenira to mesto može se očekivati u
roku od 12-18 sati
www.belimantil.info
74.
• EKSTRAĆELIJSKI MIKROORGANIZMI:
APCCD4+ T limfociti
• INTRAĆELIJSKI MIKROORGANIZMI:
APC CD8+ T limfociti
Šta se dešava kada virus inficira bilo koju ćeliju
organizma koja nije profesionalna APC ?
www.belimantil.info
75.
Ćeliju inficiranu intraćelijskimmikroorganizmom (npr. virusom) fagocituje profesionalna APĆ.
Antigeni virusa i sam virus se razgrađuju u fagocitu i prikazuju u kompleksu sa MHC molekulima
T limfocitima. Zavisno od načina prerade, razgrađeni virus ili njegovi antigeni se povezuju
u kompleks sa MHC molekulima I ili II klase. CD8+ T limfociti prepoznaju razgrađene antigene
u kompleksu sa MHC molekulima I klase, dok CD4+ T limfociti prepoznaju razgrađene antigene
u kompleksu sa MHC molekulima II klase.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanjawww.belimantil.info
MHC molekuli suproteini membrane ćelija (APC i
drugih)
Otkriće MHC molekula povezano je reakcijama
prihvatanja / odbacivanja transplantiranog tkiva
(molekuli tkivne podudarnosti, histokompatibilnosti)
Fiziološka uloga MHC molekula je “prikazivanje”
antigena u obliku koji je pogodan da bi ga T limfociti
prepoznali
MHC molekuli klase II: ekspresija na profesionalnim
APC, makrofagama, B limfocitima
MHC molekuli klase I: ekspresija na svim čelijama sa
jedrom
www.belimantil.info
78.
Geni odgovorni zastvaranje MHC molekula
organizovani su u MHC (Major
Histocompatibility Complex) genskom lookusu
koji je kod čoveka označen i kao HLA (Human
Leukocyte Antigen) lokus
MHC (HLA) lokus se sastoji iz dve grupe visoko
polimorfnih gena: klase I i klase II
Proizvodi ove dve grupe gena označeni su kao
MHC molekuli klase I i klase II
U MHC lokusu postoje i nepolimorfni geni čiji
proizvodi imaju različite funkcije
www.belimantil.info
79.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
80.
MHC geni senasleđuju kodominantno:
podjednaka ekspresija alela oba roditelja
MHC geni su visoko polimorfni: nema dve
osobe koje imaju potpuno iste grupe MHC
gena i MHC molekula
- mogućnost da svaka jedinka prikaže
gotovo neograničeni broj antigena u
sklopu MHC molekula i da na njih
razvije imuni odgovor
- problemi u transplantaciji tkiva/organa
www.belimantil.info
ŠEMATSKI PRIKAZ IMODELI KRISTALNE
STRUKTURE MHC MOLEKULA I i II KLASE
Oba molekula se sastoje iz dva polipeptidna lanca:
MHC I : a lanac i b2 mikroglobulin
MHC II : a i b lanac
Lanci MHC molekula formiraju domene:
MHC I klase :
- varijabilni domeni a1 i a2 formiraju žleb za
vezivanje antigenskog peptida veličine
8-11 amino-kiselina;
- konstantni a3 domen se vezuje za CD8
molekul naT limfocitima
MHC II klase:
- varijabilni domeni a1 i b1 formiraju žleb za
vezivanje antigenskog peptida veličine
10-30 amino-kiselina;
- konstantni b2 domen se vezuje za CD4
molekul na T limfocitima
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
83.
KAKO MHC MOLEKULVEZUJE
ANTIGEN ?
Da bi se antigen vezao za MHC
molekul, on mora da se razloži do
peptida
www.belimantil.info
84.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
Jedan MHC molekulmože da veže samo jedan peptid
tokom određenog vremena.
Peptidi se ugrađuju u žleb tokom sinteze i sklapanja
MHC molekula unutar ćelije.
Vezivanje peptida za MHC molekul je niskog afiniteta
pa jedan MHC molekul može da veže veći broj različitih
peptida u različito vreme (prilagođavanje žleba).
Vezani peptid se sporo odvaja od MHC molekula (sati-
dani), što povećava šansu T limfocitu da dođe u
kontakt sa peptidom i započne imuni odgovor.
MHC molekul mora da sadrži oba svoja lanca (a i b) i
da veže peptid da bi bio stabilan i da bi se ispoljio na
membrani ćelije; ’’prazni’’ MHC molekul se razgrađuje
u ćeliji.
MHC molekul vezuje samo peptide (strane, sopstvene)
- da li sopstveni Ag okupiraju MHC molekul ?
- zašto se ne razvija autoimuna reakcija ?
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanjawww.belimantil.info
87.
KAKO I GDESE PRERAĐUJU
PRIRODNI PROTEINSKI ANTIGENI
(MAKROMOLEKULI) DO PEPTIDA ?
www.belimantil.info
88.
Proteinski antigeni ekstraćelijskog
poreklase razlažu do peptida
u endocitnim vezikilama i povezuju
sa MHC molekulima II klase.
Kompleks peptid/MHC II prepoznaju
CD4+ T limfociti (put MHC klase II)
Proteinski antigeni intraćelijskog
porekla se razgrađuju do peptida
u proteazomima i povezuju sa
MHC molekulima I klase.
Kompleks peptid/MHC I prepoznaju
CD8+ T limfociti (put MHC klase I)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanjawww.belimantil.info
89.
Internalizacija ekstraćelijskih mikroorganizama
(fagocitoza)ili solubilih proteina (pinocitoza).
Formiranje endozomne (fagozomne) vezikule
koja se spaja sa lizozomnom vezikulom (fago-
lizozom).
Protein se razgrađuje do peptida u fagolizo-
zomu.
MHC molekul klase II se sintetiše u ER i u
žleb se umeće lanac Ii preko CLIP.
MHC II + Ii se preko egzocitne vezikule dovode
u kontakt sa peptidom.
U vezikuli se odvaja CLIP i žleb ostaje
slobodan za vezivanje peptida. Kompleks
peptid/MHC II je stabilan i putuje do membrane
gde se prikazuje CD4+ T limfocitima.
Ako se petid nije vezao za MHC II, ‘’prazan’’
MHC molekul se razgrađuje u vezikuli.
Imunodominantni epitopi = peptidi koji se
vezuju za MHC molekule
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanjawww.belimantil.info
90.
Citosolni antigeni mogupoticati od
intraćelijskih mikroorganizama (virusi,
neke bakterije) ili su proizvod fagocito-
vanih ekstraćelijskih bakterija; mogu
poticati i od izmenjenih sopstvenih gena
(npr. tumorski antigeni), ili ‘’isluženih’’
proteina ćelije.
Citosolni antigeni se razgrađuju u
proteazmima do peptida i pomoću
transportnog proteina (TAP) prenose do
ER gde se vezuju za novosintetisane
MHC molekule I klase.
Stabilan kompleks peptid/MHC I se
ispoljava na membrani ćelije gde se
prikazuje CD8+ T limfocitima. MHC
molekul I klase koji nije vezao peptid se
razgrađuje u vezikulama.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
91.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
92.
KAKAV JE FIZIOLOŠKIZNAČAJ
PRIKAZIVANJA PROTEINSKOG
ANTIGENA U SKLOPU MHC
MOLEKULA (MHC RESTRIKCIJA) ?
www.belimantil.info
93.
T limfociti ‘vide’(prepoznaju) samo peptide
koji su vezani za ćeliju (nisu solubilni) bilo da
su fagocitovani (ekstraćelijski) ili intraćelijski
antigeni
Putevi MHC klase I ili klase II za preradu
antigena omogućavaju najefikasnije
efektorske mehanizme imunog odgovora u
odbrani od ekstraćelijskih i intraćelijskih
mikroorganizama
www.belimantil.info
94.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
95.
PODSETNIK:
1. Stimulacija specifičnogimunog odgovora na proteinske antigene zavisi od posebnog
načina prihvatanja, prerade i ekspresije tog antigena u obliku u kome ga mogu
prepoznati T limfociti. Funkcije prihvatanja, prerade i ekspresije proteinskog antigena
obavljaju antigen-prezentujuće ćelije (APĆ).
2. APĆ koje se nalaze u koži i sluznicama, prihvataju antigene koji su prodrli u organizam
ovim putevima i transportuju ih do regionalnog limfnog čvora. Ako je antigen u
limfotoku, mogu ga prihvatiti i APĆ (DĆ) koje se nalaze u limfnom čvoru, a ako je
antigen u krvotoku, mogu ga prihvatiti APĆ (DĆ) koje se nalaze u slezini.
3. U perifernim limfnim organima, DĆ prikazuju proteinske antigene u formi koja je
pogodna za specifično prepoznavanje od strane “naivnih” T limfocita.
4. Pogodna forma proteinskog antigena koji prepoznaju T limfociti je: da je protein
razgrađen do peptida, da je peptid u kompleksu sa MHC molekulima i da je kompleks
peptid/MHC ispoljen na membrani APĆ (DĆ).
5. MHC molekuli su produkti MHC gena (Major Histokompatibility Compleks- geni
histokompatibilnosti ili tkivne podudarnosti). MHC molekuli I klase su ispoljeni na svim
ćelijama organizma koje imaju jedro, a MHC molekuli II klase na APĆ (DĆ, makrofagi, B
limfociti...).
6. Proteinski antigeni koji su prihvaćeni od strane APĆ iz ekstraćelijskog okruženja
(ekstraćelijski antigeni), proteolitićki se razgrađuju do peptida u vezikulama-
fagozomima ili endozomima- spajaju sa MHC molekulima II klase i prikazuju CD4+ T
limfocitima, jer se CD4 molekul vezuje za β2 domen MHC II klase.
www.belimantil.info
96.
7. Proteinski antigenikoji potiću od intraćelijskih mikroorganizama (virusi, neke bakterije),
tumorski antigeni, antigeni “isluženih” proteina, koji se razgrađuju do peptida u citosolnim
proteazomima APĆ ( intraćelijski antigeni), povezuju se sa MHC molekulima I klase i prikazuju
CD8+ T limfocitima, jer se CD8 molekul vezuje za α3 domen α lanca MHC I klase.
8. Prepoznavanje petid/MHC I kompleksa od strane CD8+T limfocita, odnosno peptid/MHC II
kompleksa od strane CD4+ T limfocita, obezbeđuje najefikasniji vid imunog odgovora protiv
intraćelijskih mikroorganizama (CD8+T limfociti CTL), odnosno ekstraćelijskih
mikroorganizama (CD4+T limfociti Th).
9. Efektorski CTL ubijaju inficirane ćelije i tumorske ćelije; efektorski Th (helper) limfociti
stimulišu makrofage da razgrade i eliminišu fagocitovane ekstraćelijske antigene i B limfocite da
sintetišu antitela značajna u odbrani od ekstraćelijskih mikroorganizama.
www.belimantil.info
97.
PREPOZNAVANJE ANTIGENA U
SPECIFIČNOM(STEČENOM)
IMUNOM ODGOVORU:
• Receptori za antigen
•Nastanak repertoara receptora u odnosu na
antigene
(repertoar specifičnosti)
www.belimantil.info
98.
RECEPTORI ZA ANTIGEN
(molekulikoji prepoznaju Ag na
limfocitima)
-kakva je struktura receptora za
Ag ?
- kako receptori prepoznaju Ag ?
www.belimantil.info
99.
Specifično prepoznavanje antigenavrše receptori za
antigen na B i T limfocitima (BCR, TCR)
BCR i TCR imaju dve funkcije:
-da otkriju (vežu) antigen
- da podstaknu odgovor limfocita za koje se vezao
antigen
BCR I TCR su klonalno raspoređeni:
jedan klon/ jedna specifičnost receptora/prepoznavanje
jednog antigena
Ukupni repertoar specifičnosti limfocita (receptora) u
odnosu na antigen je >109-1011
Iako klonovi limfocita prepoznaju različite antigene,
proces aktivacije posredstvom receptora za antigen je
uglavnom isti
www.belimantil.info
100.
OSOBINE I FUNKCIJABCR i TCR
BCR i TCR prepoznaju hemijski
različite strukture.
Raznolikost BCR i TCR je oko
109-1011 različitih antigenih
struktura.
BCR i TCR molekuli formiraju
regione/domene:
-varijabilni (V) regioni služe za
prepoznavanje antigena; u
okviru V regiona postoje
hipervarijabilni regioni ili regioni
koomplementarnosti koji su
komplementarni strukturi
antigena (kao kalup).
-konstantni (C) regioni
omogućavaju strukturni
integritet receptorskog molekula
(TCR), odnosno služe za
ostvarivanje efektorskih funkcija
(Ig).
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanjawww.belimantil.info
101.
OSOBINE I FUNKCIJABCR i TCR
BCR i TCR su nekovalentno
povezani sa molekulima
nepromenljive strukture čija je
uloga prenos aktivacionog
signala posle interakcije
Ag:receptor; zajedno ovi molekuli
čine BCR kompleks, odnosno
TCR kompleks.
TCR postoji samo kao receptor
na membrani T limfocita i nema
drugu funkciju osim
prepoznavanja antigena i
aktivacije T ćelija.
BCR su antitela (imunoglobulini)
vezani za membranu B limfocita;
antitela postoje i kao sekretorni
molekuli u telesnim tečnostima
(humorima) gde služe kao
efektorski molekuli humoralnog
imunog odgovora (eliminacija
antigena)
www.belimantil.info
102.
BCR KOMPLEKS
•Molekul antitela(imunoglobulina)
prepoznavanje antigena
•Nekovalentno vezani Igα i Igβ
prenos signala aktivacije
www.belimantil.info
103.
STRUKTURA ANTITELA
At jesastavljeno od dva identična
teška (H) lanca i dva identična laka
(L) lanca.
Na svakom od 4 lanca postoji po
jedan varijabilni (V) region i jedan
ili više konstantnih (C) regiona;
trodimenzionalna struktura regiona
čini odgovarajući domen antitela.
Veze između L i H lanaca i H i H
lanaca su disulfidne
Svaki V region H i L lanaca sadrži
po tri hipervarijabilna regiona
(regioni komplementarnosti);
najveću varijabilnost ima CDR3 koji
jse nalazi na spoju V i C regiona;
CDR3 u najvećoj meri učestvuje u
vezivanju odgovarajućeg antigena.
Proteolitičkom razgradnjom At
dobijaju se fragmenti:
-Fab fragment (Fragment antigen
binding) vezivanje Ag
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanja
STRUKTURA ANTITELA
www.belimantil.info
104.
STRUKTURA ANTITELA
-Fc fragment(Fragment
crystalline) : biološka aktivnost i
efektorske funkcije antitela
Između Fab i Fc fragmenta nalazi
se zglobni region fleksibilnost
molekula At.
C terminalni delovi H lanca
membranskih i sekretorni antitela
se razlikuju: membranska imaju
deo lanca za ugradnju u
membranu B limfocita, a
sekretorna imaju kratak “rep”
Postoji dva tipa L lanaca: κ i λ .
Postoji pet tipova H lanaca: γ, α, μ,
δ, ε na osnovu kojih se antitela
dele na izotipove ili klase IgG, IgA,
IgM, IgD, IgE.
Različiti izotipovi (klase) antitela
imaju različite fizičko-hemijske
osobine i različite biološke uloge
www.belimantil.info
105.
Izotip/ Podklasa AtH lanac Koncentracija Poluživot Sekretorna Funkcije
Klasa At u serumu u serumu forma
(mg/ml) (dani)
IgA A1,A2 α1, α2 3.5 6 monomer Imunitet sluzica,
dimer mukusa, neonatalni
trimer pasivni imunitet
IgD nema δ tragovi 3 nema Receptor za Ag
na B limfocitima
IgE nema ε 0.05 2 monomer Reakcije rane
preosetljivosti
IgG IgG1-G4 γ1−γ4 13.5 23 monomer Opsonizacija,
aktivacija komplemen.
ADCC, neonatalni
pasivni imunitet,
regulac. imun.odgov.
IgM nema μ 1.5 5 pentamer Receptor za Ag
na B limfocitima,
aktivacija komplemen.
www.belimantil.info
Region za vezivanjeantigena (paratop) formira ravnu površinu sposobnu da se prilagodi različitim
oblicima antigena;
Veze Ag/At su reverzibilne, nekovalentne (vodonične, jonske).
Delovi antigena koje prepoznaju antitela zovu se epitopi (antigenske determinante); epitopi mogu biti
linearni (sekvence), konformacioni (trodimenzionalni) ili neoepitopi (nastali pod uticajem fizičkih ili
hemijskih agenasa).
Jačina veze između jednog mesta na antitelu koje vezuje antigen (paratop) i jednog epitopa antigena je
afinitet.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
iskljućivo za potrebe predavanja
www.belimantil.info
108.
Afinitet se izražavai kao konstanta disocijacije Kd; Kd je molarna koncentracija antigena potrebna da
se veže za polovinu molekula antitela prisutnih u nekom rastvoru što je manja Kd, veći je afinitet.
Afinitet antitela je 106-109M, ali se on povećava do 108-1011M u sekundarnom imunom odgovoru
afinitetno sazrevanje.
Antitelo (monomer, dimer, trimer, pentamer) može da veže 2-10 jednakih epitopa na antigenu pod
uslovom da su epitopi dovoljno blizu jedan drugom. Ukupna jačina vezivanja svih paratopa i epitopa je
aviditet.
Antitela stvorena protiv jednog antigena mogu da reaguju i sa drugim strukturno sličnim antigenom:
ukrštena reaktivnost.
www.belimantil.info
109.
TCR KOMPLEKS
•TCR molekul
prepoznavanjeantigena
•CD3 molekul + ζ (ceta) lanci
prenos signala aktivacije
•Koreceptorski molekuli CD4 i
CD8
www.belimantil.info
110.
Heterodimer (α iβ lanac).
Oba lanca imaju varijabilni (V) i
konstantni (C) region.
Vα i Vβ sadrže po tri hipervarijabilna
dela (regioni komplementarnosti),
treći je najvarijabilniji.
TCR se ne stvara u sekretornoj formi
i ne podleže afinitetnom sazrevanju.
Oba lanca (α i β) imaju ekstraćelijski
deo, transmembranski i kratki
citoplazmatski deo.
Preko 90% T limfocita ima TCRαβ, a
oko 10% TCRγδ. TCRγδ+T limfociti su
u najvećem broju prisutni u epitelu.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanja
STRUKTURA TCR
www.belimantil.info
TCR prepoznaje samo1-3
rezidue peptida vezanog za
MHC molekul.
TCR prepoznaje samo nekoliko
peptida složenog
makromolekula:
imunodominantni epitopi.
TCR prepoznaje istovremeno i
polimorfne delove sopstvenog
MHC molekula koji nosi peptid.
U prepoznavanju peptid/MHC
kompleks ućestvuju Vα i Vβ
domeni TCR
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe
predavanjawww.belimantil.info
113.
Shema preuzeta izknjige Basic Immunology,
Abbas i Lichtman, iskljućivo za potrebe
predavanja
Sličnosti i razlike prepoznavanja antigena
od strane BCR i TCR
www.belimantil.info
NASTANAK I SAZREVANJELIMFOCITA
Limfociti nastaju iz neopredeljenih matičnih ćelija kostne
srži kao i druge krvne ćelije
Proces nastanka limfocita uključuje:
- proliferaciju nezrelih ćelija prekursora
- ekspresiju (ispoljavanje) receptora za antigen
- selekciju limfocita koji ispoljavaju “korisne” receptore
za antigen
Sazrevanje B limfocita se u potpunosti odvija u kostnoj
srži, a T limfocita u timusu
www.belimantil.info
116.
1. PROLIFERACIJA: signaliza proliferaciju i preživljavanje preko IL7R i intaktnih TCR ili BCR
Shema preuzeta iz knjige
Basic Immunology, Abbas i
Lichtman, iskljućivo za
potrebe predavanja
2. EKSPRESIJA TCR ili BCR:TCR i BCR su proizvodi nekoliko genskih lokusa/segmenata koji se rekombinuju
(preuređuju) tokom sazrevanja limfocita dajući receptore različitih specifičnosti za antigen. Ekspresija TCR i BCR
različitih specifičnosti je najvažnije zbivanje tokom sazrevanja limfocita.
3. SELEKCIJA: U toku sazrevanja limfociti podležu selekciji (odabiru) kako bi se sačuvale korisne osobine.
Pre-limfociti koji ne ispolje pre-receptor umiru.
Nezreli T limfociti se odabiru tako da prepoznaju sopstvene MHC molekule (pozitivna selekcija).
Nezreli T i B limfociti se takođe odabiru tako da ne prepoznaju sopstvene antigene visokim afinitetom (negativna
selekcija) čime se uklanjaju potencijalno štetni autoreaktivni limfociti.
www.belimantil.info
117.
Najvažniji događaj tokomsazrevanja
limfocta je rekombinacija
(preuređivanje) gena i ekspresija
receptora za antigen
www.belimantil.info
118.
Kako nastaje različitost(>109) receptora
za antigen (repertoar specifičnosti) ?
Repertoar se formira nasumičnim kombinacijama
gena za antigenske receptore i nevezano sa
prisustvom antigena
Nastanak repertoara omogućen je
karakterističnom organizacijom i rekombinacijom
gena za antigeske receptore
www.belimantil.info
119.
Genski lokusi strukturnih
komponentiBCR i TCR nalaze se na
različitim hromozomima.
U genomu matičnih ćelija lokusi su
nepreuređeni (nearanžirani): svaki
lokus sadrži veći broj (nekoliko
stotina) gena varijabilnog (V) i jedan
ili nekoliko gena konstantnog (C)
regiona.Između V i C gena nalazi se
nekoliko malih segmenata: J (joining,
spajanje) i D (diversity, raznolikost).
D segment se nalazi samo na lokusu
teškog (H) lanca Ig i lokusu β TCR.
Usmeravanje sazrevanja progenitora
ka limfocitima ( pro-limfociti)
uključuje nasumičnu, slučajnu
rekombinaciju jednog V gena sa
jednim D i jednim J genom.: V-D-J
rekombinacija. Rekombinacije se
dešavaju pod uticajem V(D)J
rekombinaza.
Preuređen V-D-J kompleks povezuje
se sa najbližim C genom i formira se
V-D-J-C mRNK odgovorna za sintezu
jednog lanca receptora za Ag
(stadijum pre-limfocita).
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
iskljućivo za potrebe predavanja
www.belimantil.info
120.
Zatim počinju dase preuređuju geni za
drugi lanac receptora za antigen po istom
principu. Sintetiše se kompletan
receptorski molekul BCR ili TCR i
ispoljava na membrani B, odnosno T
limfocita (stadijum nezrelih limfocita)
* * * *
Raznolikost secifičnosti receptora
za antigen (>109) rezultat je:
• raznolikosti kombinacija V-D-J
segmenata gena (zavisi od broja
gena u segmentima) i
• raznolikosti povezivanja V-D, D-J
www.belimantil.info
121.
1. Sazrevanje Blimfocita odvija se u celini u kostnoj srži. Neopredeljena matična ćelija pod
uticajem IL-7 daje veliki broj pro-B ćelija. Geni za BCR su nearanžirani u ovim ćelijama, mada
u pro-B ćelijama započinje rearanžman V-D-J gena teških (H) lanaca.
2. Iz pro-B ćelija nastaju pre-B ćelije u kojima je došlo do preuređenja gena za Hμ lanac Ig.
Μ lanac se nalazi u citoplazmi pre-B ćelija ili se ispoljava na membrani zajedno sa
nepreuređenim (surogatnim) L lancem: pre-B receptor za antigen. Ovaj receptor omogućava
preživljavanje i proliferaciju ćelija, a ćelije koje ne ispolje pre-B receptor umiru prva
kontrola. Signali preko pre-B receptora utiču na zaustavljanje rekombinacije gena H lanca na
drugom hromozomu (isključivanje alela) i podsticanje rekombinacije gena κ ili λ L lanaca.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanja
SAZREVANJE I SELEKCIJA B LIMFOCITA
www.belimantil.info
122.
SAZREVANJE I SELEKCIJAB LIMFOCITA
3. Iz pre-B ćelija nastaju nezrele B ćelije koje stvaraju kompletan membranski IgM receptor (BCR)
(Hμ + Lκ ili Lλ). Ovaj receptor ponovo omogućava preživljavanje i proliferaciju ćelija i zaustavlja
rekombinaciju gena drugog L lokusa (jedna B ćelija i njeni potomci stvaraju samo κ ili samo λ
lake lance) druga kontrola. Ako nezreli B limfocit veže pomoću mIgM (BCR) antigen
visokim afinitetom, zaustavlja se dalje sazrevanje ove ćelije (negativna selekcija). Ćelija ili
umire apoptozom ili reaktivira enzime jedra da preurede drugi L lanac i tako izmene
specifičnost BCR (reprogramiranje receptora). Procesom negativne selekcije se uklanjaju
potencijalno štetne autoreaktivne B ćelije.
4. Iz nezrelih B ćelija nastaju zreli B limfociti na čijoj membrani su ispoljeni istovremeno IgM i IgD
BCR. Sinteza druge klase imunoglobulina iste specifičnosti za antigen omogućena je
alternativim spajanjem prethodno rekombinovanog V-D-J segmenta sa Cδ na nivou mRNK.
Samo B limfociti koji istovremeno ispoljavaju IgM i IgD receptore su sposobi da odgovore na
antigen.
www.belimantil.info
123.
SAZREVANJE I SELEKCIJAT LIMFOCITA
1. Iz neopredeljenih matičnih ćelija kosne srži nastaju opredeljene ćelije T limfocitne loze, pro-
T ćelije, koje su “dvostruko negativne” CD4-CD8-. Pro-T ćelije migriraju iz kosne srži u timus gde
se odvija kompletno sazrevanje T limfocita. Pod dejstvom IL-7 ove ćelije proliferišu i u njima se
dešava V-D-J rekombinacija gena TCRβ lokusa.
2. Na pre-T ćelijama se ispoljava β lanac TCR zajedno sa nearanžiranim α lancem: pre-TCR.
Preko pre-TCR se prenose signali koji obezbeđuju preživljavanje i proliferaciju ćelija, isključivanje
alela TCRβ na drugom hromozomu i rearanžman gena TCRα lokusa. Pre-T ćelije koje ne ispolje
kompletan β lanac TCR umiru.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanja
SAZREVANJE I SELEKCIJA T LIMFOCITA
www.belimantil.info
124.
SAZREVANJE I SELEKCIJAT LIMFOCITA
3. Iz pre-T ćelija nastaju nezreli T limfociti koji su “dvostruko pozitivni” CD4+CD8+ i koji
ispoljavaju kompletan TCRαβ. Nezreli T limfociti koji u timusu prepoznaju niskim afinitetom
sopstvene peptide u kompleksu sa sopstvenim MHC molekulima klase I ili II preživljavaju
(pozitivna selekcija), dok limfociti koji visokim afinitetom prepoznaju sopstveni
peptid/sopstveni MHC kompleks umiru apoptozom (negativna selekcija). Negativnom
selekcijom se uklanjaju potencijalno štetni autoreaktivni T limfociti.
4. U toku procesa pozitivne selekcije nezreli T limfociti koji niskim afinitetom prepoznaju
sopstveni peptid/sopstveni MHC klase I gube CD4 molekul, a limfociti koji prepoznaju
sopstveni peptid/MHC klase II gube CD8 molekul. Tako nastaju zreli T limfociti koji su
“jednostruko pozitivni” ili CD4+ ili CD8+. Ove dve populacije limfocita su funkcionalno
različite.
www.belimantil.info
125.
Imuni odgovor
Urođeni (prirodni,nespecifični) imunitet
Stečeni (adaptivni, specifični) imunitet:
humoralni
celularni
aktivni, pasivni (prirodni, veštački)
Faze imunog odgovora: prepoznavanje, aktivacija, efektorska faza, zaustavljanje
Ćelije imunog sistema: limfociti, APC(DC), NK, makrofagi, neutrofili
Organi: centralni (primarni, generativni), periferni (sekundarni)
Tkiva: GALT, BALT, difuzna
Specifični imuni odgovor na proteinske antigene započinje interakcijom:
Ag peptida
MHC molekula
TcR
www.belimantil.info
126.
Transport, obrada, prezentacijaproteinskih antigena:
ekstraćelijski antigeni
intraćelijski antigeni
Ag peptid/MHC I...................CD8+ T limfociti
Ag peptid/MHCII...................CD4+ T limfociti
Struktura i distribucija MHC I i MHC II
Receptori za antigen BcR i TcR ( BcR i TcR kompleks)
BcR i TcR prepoznaju različite hemijske prirode i oblike antigena
BcR: membranski imunoglobulin (antitelo)
- struktura antitela
- funkcionalni domeni At
- klase At
- interakcija Ag (epitopa) i At (paratopa)
TcR: - struktura
- funkcionalni domeni
- interakcija sa Ag peptid/MHC kompleksom
Nastanak repertoara specifičnosti (antigenski repertoar)
- struktura i preuređenje (rearanžman) gena za BcR i TcR
- mehanizmi raznolikosti
- selekcija limfocita
www.belimantil.info
127.
PODSETNIK:
1. Specifićno prepozavanjeantigena vrše receptori za antigen na T i B limfocitima
(TCR i BCR)
2. TCR i BCR imaju dve funkcije: da vežu antigen i da započnu aktivaciju limfocita
3. TCR i BCR su klonalno raspoređeni
4. BCR predstavlja membransku formu imunoglobulina (antitela)
5. TCR je heterodimer (α/β ili u manjem procentu γ/δ)
6. Varijabilni regioni TCR i BCR služe za prepoznavanje antigena, a konstantni regioni
čuvaju strukturni integritet TCR, odnosno služe za ostvarivanje brojnih
efektorskih funkcija antitela
7. TCR postoji samo kao receptor na membrani T limfocita; BCR postoji kao
membranska forma antitela (imunoglobulina) i kao sekretorni molekul u telesnim
tečnostima
8. BCR kompleks se sastoji iz molekula antitela (prepoznavanje antigena) i
nekovalentno vezanih Igα i Igβ (prenos signala aktivacije)
9. TCR kompleks se sastoji iz TCR (prepoznavanje antigena) i nekovalentno povezanih
CD3 molekula i ζ lanaca (prenos signala aktivacije); koreceptorski molekuli CD4 i
CD8
www.belimantil.info
128.
10. Struktura antitelai TCR
11. Klase antitela i njihova funkcija
12. Vezivanje antitela za antigen: paratopi, epitopi; afinitet i aviditet antitela
13. Raznolikost TCR i BCR (repertoar specifičnosti, antigenski repertoar 109- 1011)
omogućena je jedinstvenom organizacijom i načinom preuređenja
(rekombinacije) gena za TCR i BCR tokom sazrevanja limfocita; procesi
preuređenja gena dešavaju se nezavisno od uticaja antigena
14. Sazrevanje limfocita uključuje:
proliferaciju ćelija prekursora
ispoljavanje receptora za antigen
selekciju (pozitivnu i negativnu) limfocita koji ispoljavaju “korisne” receptore
www.belimantil.info
129.
IMUNI ODGOVOR TLIMFOCITA-
CELULARNI IMUNI ODGOVOR:
Nastanak efektorskih T limfocita
Faze :
1. Prepoznavanje antigena
2. Aktivacija T limfocita
3. Proliferacija T limfocita (ekspanzija klona)
4. Diferencijacija naivnih T limfocita u
efektorske ćelije
www.belimantil.info
130.
Suština T-ćelijskog (celularnog)imunog
odgovora je:
1. Odbrana organizma od infekcija intracelularnim
mikroorganizmima (neke bakterije, gljivice, protozoe, virusi)
koji su:
fagocitovani, ali su rezistentni prema proteolitičkim
enzimima fagolizozoma;
izašli iz fagozoma i preživljavaju/umnožavaju se u
citoplazmi inficiranih ćelija;
sposobni da se umnožavaju u citoplazmi inficirane ćelije
ili se njihova DNK integriše u genom ćelije domaćina (virusi)
2. Pomoć B limfocitima da stvore antitela
T-odgovor
www.belimantil.info
131.
T- odgovor
A. Nekifagocitovani
(ekstraćelijski)
mikroorganizmi mogu
preživljavati u
fagolizozomima jer su
rezistentni na lizozomne
enzime, ili mogu izaći iz
vezikula u citoplazmu gde
takođe preživljavaju.
B. Virusi mogu inficirati
rezličite ćelije i
razmnožavati se u njihovoj
citoplazmi; neki virusi
izazivaju latentne infekcije
u kojima inficirane ćelije
proizvode proteine virusa
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
isključivo za potrebe predavanja
www.belimantil.info
132.
FazeA
“Naivni” T limfocitiprepoznaju
antigeni peptid/MHC kompleks
ispoljen na membrani APC (prvi
signal aktivacije). Za potpunu
aktivaciju T limfocita potrebni su i
dodatni kostimulacioni signali koji
obezbeđuju drugi signal aktivacije.
Istovremeno sa prepoznavanjem
antigena, “naivni” T limfociti
primaju i signale koji potiču od
samog mikroorganizma ili
komponenti nespecifičnog imunog
odgovora na infekciju. Rezultat
ovih stimulacija je da “naivni” T
limfociti počinju da proizvode
citokine koji stimulišu njihovu
proliferaciju. Umnožavanje T
limfocita specifičnih za dati
antigeni peptid je klonalna
ekspanzija.
Deo aktivisanih T limfocita se
transformiše (diferencijacija) od
“naivnih” u efektorske T ćelije, a
deo u memorijske T ćelije.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanjawww.belimantil.info
133.
FazeB
Procesi prepoznavanja antigena,
aktivacije,klonalne ekspanzije i
nastanka efektorskih T limfocita
odvijaju se u perifernom
limfoidnom tkivu.
Efektorske T ćelije napuštaju
periferno limfoidno tkivo i migriraju
do bilo kog mesta u organizmu koje
je inficirano, gde uklanjaju
mikroorganizam. Nakon obavljene
funkcije, efektorske ćelije umiru, a
imuni sistem se vraća u stanje
mirovanja (imuni odgovor se
prekida).
Deo aktivisanih T limfocita se
razvija u memorijske T ćelije koje
dugo žive (meseci, godine),
funkcionalno su neaktivne i
neprestano recirkulišu kroz
organizam spremne da brzo
reaguju pri ponovnom kontaktu
organizma sa istim
mikroorganizmom.
www.belimantil.info
134.
Aktivacija
Početak T-ćelijskog odgovora
zahtevavišestruke interakcije
različitih molekula na
membranama T limfocita i APC.
Svi ovi molekuli, izuzev TCR,
zajednički se zovu akcesorni
molekuli.
U funkcionalnom smislu,
akcesorni molekuli se mogu
podeliti u 3 kategorije:
- Koreceptorski molekuli: CD4,
CD8;
- Molekuli za prenos signala:
CD3, ζ lanci, CD4, CD8, CD28,
CTLA-4:
- Adhezioni molekuli: CD4, CD8,
LFA-1, VLA-4
Akcesorni molekuli na T
limfocitima reaguju sa
odgovarajućim ligandima na
APC
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanjawww.belimantil.info
135.
Aktivacija
TCR i koreceptori(CD4 ili
CD8) zajedno prepoznaju
peptid/MHC kompleks na
APC prvi signal
aktivacije.
TCR prepoznaje peptid i
rezidue MHC molekula
koje su oko žleba za
vezivanje peptida, dok
CD4, odnosno CD8
molekuli prepoznaju
mesta na MHC molekulu
klase II, odnosno klase I
koja su izvan žleba za
smeštanje peptida.
Potrebno je da se dva ili
više TCR i koreceptora
(CD4 ili CD8) istovremeno
poveže sa peptid/MHC
kompleksom kako bi se
pokrenula aktivaciona
kaskada.
Funkciju prenosa prvog
signala aktivacije imaju
CD3 i ζ lanci (ulaze u
sastav TCR kompleksa) i
koreceptori CD4 ili CD8.
KORECEPTORSKI
MOLEKULI
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
136.
Aktivacija
ADHEZIONI MOLEKULI
Interakcija adhezionihmolekula na T limfocitima sa odgovarajućim ligandima na
APC ima za cilj da stabilizuje, učvrsti vezu T ćelija: APC i time omogući
uspostavljanje aktivacionih signala.
Najznačajniji adhezioni molekuli pripadaju grupi heterodimernih proteina –
integrina. Integrin LFA-1 (limfocitni funkcioni antigen-1), koji je ispoljen na T
limfocitima, reaguje sa ICAM-1 (intercelularni adhezioni molekul-1), koji je
ispoljen na APC.
Integrini su značajni i za usmereno kretanje efektorskih T ćelija iz cirkulacije do
mesta infekcije
www.belimantil.info
137.
Aktivacija
KOSTIMULACIONI MOLEKULI
Za potpunuaktivaciju T limfocita neophodna su dva signala: prvi signal se ostvaruje posredstvom TCR kompleksa/koreceptora, a
drugi signal posredstvom kostimulacionih molekula.
Prvu grupu kostimulatora čine CD28 molekul na T limfocitima i ligandi B7-1 (CD80) i B7-2 (CD86) na APC.
Drugu grupu kostimulatora čine CD40L (ligand) na T limfocitima i CD40 molekul na APC. Interakcija CD40:CD40L ne
aktiviše direktno T limfocit, već stimuliše APC da ispolji više B7 molekula i da sekretuje citokine
Ako izostane interakcija CD28:B7, signal preko TCR nije dovoljan za aktivaciju T ćelija, čak može da nastane dugotrajna tolerancija
(nereaktivnost) antigen-specifičnih T ćelija.
Shema preuzeta iz knjige Basic Immunology,
Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
138.
Aktivacija
KOSTIMULACIONI MOLEKULI
I zapotpunu aktivaciju CD8+
T limfocita neophodan je
drugi signal aktivacije koji
obezbeđuju kostimulatori.
Ako je kompleks peptid/MHC I
ispoljen na membrani APC,
ove ćelije obezbeđuju i
kostimulaciju T limfocita.
Ako je kompleks peptid/MHC I
ispoljen na membrani bilo
koje ćelije u organizmu, takve
ćelije ne obezbežuju
kostimulaciju T limfocita. U
tom slučaju, CD4+T limfociti
(Th ili pomoćnici) iz okoline
inficirane ćelije mogu da
stvaraju citokine ili druge
molekule preko kojih
obavljaju kostimulaciju
antigen-specifičnih CD8+ T
limfocita.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanjawww.belimantil.info
139.
Aktivacija
Značaj kostimulacije:
Bez kostimulacijenema T-ćelijskog odgovora
Vakcine koje sadrže proteinske antigene moraju
da sadrže i adjuvanse (supstance koje indukuju
ekspresiju kostimulatora na APC i sekreciju citokina)
Stimulacija ekspresije kostimulatora je značajna u
savremenim terapijskim strategijama tumora
Blokiranje ekspresije i funkcije kostimulatora je
značajno u savremenim terapijskim strategijama
autoimunih bolesti i odbacivanja transplantiranih organa
www.belimantil.info
140.
AktivacijaA
BIOHEMIJSKI PUTEVI SIGNALA
AKTIVACIJET LIMFOCITA
Prilikom prepoznavanja kompleksa
peptid/MHC, veći broj TCR i koreceptora
(CD4 ili CD8) se grupiše na membrani T
limfocita. Grupisanje CD4 ili CD8 dovodi
do ativacije enzima protein tirozin kinaze
Lck.
Lck fosforiliše tirozinske rezidue ITAM
(imunoreceptor tirozinske aktivacione
sekvence) citoplazmatskih delova CD3 i ζ
lanaca TCR kompleksa.
Fosforilisane ITAM sekvence z lanaca
vezuju tirozin kinazu ZAP-70 (ζ
associated protein-70kD).
Aktivisani ZAP-70 aktiviše brojne
adapterske proteine i enzime koji
prenose signal aktivacije. Dva najvažnija
puta prenosa signala su:
•Ca-NFAT put (nuklearni faktor
aktivisanih T)
•Ras-MAP put
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
isključivo za potrebe predavanja
www.belimantil.info
141.
AktivacijaB
BIOHEMIJSKI PUTEVI SIGNALA
AKTIVACIJET LIMFOCITA
Ca-NFAT put započinje tako što ZAP-70
vrši aktivaciju PLC (fosfolipaza C). PLC
deluje na membranske IP (inozitol
fosfolipidi) cepajući ih na IP3 (inozitol
trifosfat) i DAG (diacilglicerol.
- IP3 stimuliše porast Ca++ jona koji
se vezuju za protein kalmodulin.
Ca++kalmodulin aktiviše kalcineurin, a
ovaj vrši defosforilaciju citosolnog
transkripcionog faktora NFAT.
Defosforilisani NFAT migrira u jedro gde
aktiviše promotore gena za IL-2 i IL-2R
(lek ciklosporin inhibira aktivnost
kalcineurina i tako sprečava produkciju
citokina; primena leka u transplantaciji i
autoimunim bolestima)
- DAG aktiviše PKC (protein kinaza
C), koja aktiviše NF-kB (nuklearni faktor-
κB); aktivisani NF-κB odlazi u jedro gde
aktiviše transkripciju nekoliko gena.
www.belimantil.info
142.
AktivacijaC
BIOHEMIJSKI PUTEVI SIGNALA
AKTIVACIJET LIMFOCITA
Ras-MAP kinaza put uključuje Ras
protein koji postaje biološki aktivan kada
se veže za GTP (guanozin trifosfat),
nekoliko adapter proteina i kaskadu
enzima nazvanih MAP kinaze (mitogen
aktivacioni protein).
Aktivisane MAP kinaze fosforilišu protein
c-Jun. Vezivanjem c-Jun za c-Fos
omogućena je aktivacija transkripcionog
faktora AP-1 (aktivacioni protein-1) koji
stimuliše transkripciju nekoliko T-
ćelijskih gena.
www.belimantil.info
143.
Odgovor T limfocitana stimulaciju
antigenom (prvi signal) i kostimulatorima
(drugi signal) je:
Sinteza i sekrecija različitih citokina
Proliferacija T limfocita specifičnih za taj
antigen (klonalna ekspanzija)
Diferencijacija antigen-specifičnih T
limfocita u efektorske ili memorijske T ćelije
www.belimantil.info
144.
OPŠTE OSOBINE CITOKINA
OsobinaMehanizam
Proizvode se povremeno kao odgovor na TCR signal i kostimulacija indukuju transkripciju
antigen gena
Obično deluju na istu ćeliju koja ih je Posle aktivacije T ćelija, dolazi do ekspresije i
produkovala (autokrino) ili na okolne citokina i njihovih receptora
ćelije (parakrino)
Pleiotropizam: jedan citokin ima različite Različite ćelije ispoljavaju receptore za jedan
biološke efekte citokin
Različiti citokini mogu imati isti ili sličan Različiti citokini korste iste puteve za prenos
biološki efekat signala
BIOLOŠKI EFEKTI NEKIH T-ĆELIJSKIH CITOKINA
Citokin Najvažniji efekat Ćelije koje ga stvaraju
IL-2 Stimuliše proliferaciju T ćelija CD4+ i CD8+ T ćelije
IL-4 Sinteza IgE CD4+ T ćelije, mast ćelije
IL-5 Aktivacija eozinofila CD4+ T ćelije, mast ćelije
IFN−γ Aktivacija makrofaga CD4+ i CD8+T ćelije, NK
TGF-β Inhibicija aktivacije T ćelija CD4+ T ćelije i razne druge
ćelije
A
B
www.belimantil.info
145.
Proliferacija
Prvi citokin kojistvaraju
aktivisani CD4+T limfociti
je IL-2 (1-2 sata posle
aktivacije); istovremeno
sa sintezom IL-2, sintetiše
se i treći lanac IL-2R, čime
ovaj receptr stiče visoki
afinitet vezivanja liganda,
tj. IL-2.
IL-2 deluje pre svega na T
limfocit koji ga je stvorio
(autokrino) podstičući
deobu ćelije (IL-2 se još
zove i T-ćelijski faktor
rasta).
CD8+T limfociti koji su
prepoznali antigen,
nemaju sposobnost da
sekretuju velike količine
IL-2; zato IL-2 koji
sekretuju antigen-
specifične CD4+T ćelije
može da izazove i
proliferaciju antigen-
specifičnih CD8+T ćelija
(parakrino dejstvo)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanjawww.belimantil.info
146.
Proliferacija
1-2 dana posleaktivacije, T limfociti počinju da proliferišu; rezultat je
umnožavanje T limfocita specifičnih za antigen koji ih je aktivisao:
klonalna ekspanzija.
Proliferacija antigen-specifičnh T limfocita posle aktivacije je intenzivna.
Na primer, broj “naivnih” CD8+T limfocita specifičnih za neki antigen
virusa je 1 na 105-106 svih limfocita u organizmu; nedelju dana nakon
infekcije tim virusom, broj antigen-specifičnih CD8+T limfocita dostiže 10-
20% svih limfocita (povećanje broja preko 10000 puta, ili na svakih 6 sati
broj ovih ćelija se udvostručuje). Antigen-specifični CD4+T limfociti se
umnožavaju u nešto manjoj meri (100-1000 puta). Razlog za različiti nivo
klonalne ekspanzije CD4+ i CD8+T limfocita verovatno leži u različitoj
funkciji ovih ćelija: CD8+ T limfociti diferenciraju u citotoksične T
limfocite (CTL) koje imaju direktno efektorsko dejstvo (ubijaju inficiranu
ćeliju), dok CD4+ T limfociti diferenciraju u ćelije koje sekretuju citokine i
posredstvom ovih molekula aktiviraju druge efektorske ćelije da uklone
mikroorganizam (npr., makrofage).
Prilikom infekcije mikroorganizmom koji sadrži veći broj antigenih
molekula, broj antigen-specifičnih klonova koji se umnožava je 1-5, tj.
znatno manji nego što taj mikroorganizam sadrži antigena. Ovi klonovi
limfocita su specifični prema imunodominantnim peptidima tog
mikroorganizma.
www.belimantil.info
147.
Efektorske ćelije
Uporedo saklonalnom ekspanzijom, dešava se i diferencijacija antigen-
specifičnih T ćelija u efektorske T ćelije; prve efektorske ćelije se
javljaju 3-4 dana od infekcije.
Diferencijacija u efektorske ćelije podrazumeva sticanje sposobnosti T
limfocita da sintetišu različite citokine (Th i CTL), ili proteolitičke
proteine (CTL).
Efektorske ćelije napuštaju periferne limfoidne organe i upućuju se na
mesto infekcije. Tamo ponovo prepoznaju antigen koji je stimulisao
njihov nastanak i brzo reaguju kako bi taj antigen eliminisale.
www.belimantil.info
148.
Efektorske ćelije/CD4+
CD4+Th ćelije
odgovarajuna antigen
tako što ispoljavaju
neke molekule na
membrani i proizvode
sekretorne molekule-
citokine.
Najvažniji membranski
molekul je CD40L. Ovaj
molekul se vezuje za
CD40 molekul koji je
ispoljen na membrani
APC (makrofage, DC, B
limfociti); rezultat
interakcije CD40L:CD40
je aktivacija APC.
Aktivisane makrofage
ubijaju fagocitovane
mikroorganizme, dok
aktivisani B limfociti
sekretuju antitela koja
neutrališu i eliminišu
antigen.
Zašto CD4+Th limfociti
u nekim slučajevima
aktivišu makrofage, a u
drugim B limfocite?
www.belimantil.info
149.
Efektorske ćelije/CD4+
Tokom diferencijacije
CD4+Tćelija u efektorske
Th ćelije, mogu nastati
dve podgrupe Th
ćelija: Th1 i Th2,
koje stvaraju različite
citokine i imaju različite
efektorske funkcije.
Th1 ćelije stvaraju IFN-
γ i stimulišu ubijanje
mikroorganizama putem
fagocitoze
Th2 ćelije stvaraju IL-4
koji stimuliše stvaranje
IgE antitela i IL-5 koji
aktivira eozinofile, što je
značajno za ubijanje
mikroorganizama
(helminti) posredstvom
eozinofilnih granulocita
Citokini porekla Th1 ćelija
imaju antagonističko
dejstvo u odnosu na
citokine Th2 ćelija i
brnuto:
Th1/Th2 dihotomija
www.belimantil.info
150.
Efektorske ćelije/CD4+
U kompravcu će se diferencirati
Th ćelije (Th1 ili Th2) tavisi od
vrste stimulusa koje tokom
diferencijacije prima CD4+T ćelija:
Ako na aktivisane CD4+T
limfocite deluje IL-12 (stvaraju
ga APC u odgovoru na
mikroorganizme (bakterije ili
viruse), nastaće Th1 efektorske
ćelije.
Ako APC ne stvaraju IL-12 u
odgovoru na mikroorganizme
(helminti), tada T ćelije (ili neke
druge ćelije?) same stvaraju IL-4
i usmeravaju diferencijaciju u
pravcu Th2 efektorskih ćelija.
www.belimantil.info
151.
Efektorske ćelije/CD8+
CD8+ Tlimfociti aktivisani antigenom i kostimulatorima ili
citokinima koje proizvode Th ćelije diferenciraju se u
citotoksične T limfocite (CTL).
CTL direktno ubijaju inficiranu ćeliju koja nosi taj antigen.
CTL ubijaju inficirane ćelije tako što luče proteine koji prave
pore na membrani i indukuju fragmentaciju DNK inficirane ćelije.
Inficirana ćelija umire putem programirane ćelijske smrti
(apoptoze)
www.belimantil.info
152.
EFEKTORSKI MEHANIZMI ĆELIJSKOGIMUNITETA
Specijalizovani imuni mehanizmi čija je funkcija
eradikacija intracelularnih mikroorganizmama
Posredovani efektorskim T-limfocitima
Antitela ne mogu eliminisati intracelularne
mikroorganizme
?
Kako efektorski T-limfociti pronalaze intracelularne
mikroorganizme na bilo kom mestu u organizmu ?
Kako efektorski T-limfociti eliminišu infekcije izazvane
intracelularnim mikroorganizmima ?
www.belimantil.info
153.
Ćelijski imunitet prema
intracelularnimmikroorganizmima
• U eliminaciji različitih tipova intracelularnih
mikroorganizmama učestvuje dva tipa
ćelijskog imuniteta
• CD4+ efektorski T-limfociti (TH1
subpopulacija) prepoznaju antigene
mikroorganizama koji su endocitovani od
strane fagocita
• Aktivirani CD4+ efektorski T-limfociti
sekretuju citokine koji aktiviraju makrofage i
stimulišu inflamaciju
• Aktivirani makrofagi ubijaju fagocitovane
mikroorganizme
• CD8+ efektorski T-limfociti prepoznaju
antigene na inficiranim ćelijama.
• Aktivisani CD8+ efektorski T-limfociti ubijaju
inficirane ćelije
• Antigeni intracelularnih mikroorganizma se
mogu naći u vezikulama (egzogeni put
prerade i asocijacija sa MHC molekulima II
klase) ili u citoplazmi (endogeni put prerade i
asocijacija sa MHC molekulima I klase)
• Patogeni mikroorganizmi koji inficiraju i
preživljavaju u ćelijama domaćina su:
1) brojne bakterije i neke protozoe koje
nakon ingestije preživljavaju u vezikulama
izbegavajući fuziju sa lizozomima ili
delovanje proteolitičkih supstanci;
2) virusi koji inficiraju fagocitne i nefagocitne
ćelije i preživljavaju u citoplazmi zaraženih
ćelija
www.belimantil.info
154.
Migracija naivnih iefektorskih T-limfocita
• Naivni T-limfociti migriraju iz krvi u
limfni čvor posredstvom
postkapilarnih venula HEV-a
• Migracija (“homing”) se odvija
interakcijom L-selektina (CD62L) na
naivnim T-limfocitima sa
odgovarajućim ligandima
(vaskularni adresini) na endotelnim
ćelijama HEV-a
• Aktivisani T-limfociti (uključujući i
efektorske T-limfocite) napuštaju
limfni čvor preko eferentnih
limfatičkih sudova, a zatim preko
ductus thoracicus-a odlaze u krv; Iz
krvi u periferna tkiva (mesta
infekcije, inflamacije)
• Aktivisani T-limfociti i efektorski T-
limfociti ispoljavaju adhezivne
molekule (ligand za E- ili P- selektin,
aktivisanu formu LFA-1 i VLA-4),
koji im omogućavaju migraciju u
periferna tkiva. Istovremeno oni
gube L-selektin zbog čega im je
onemugućena ponovna migracija
(recirkulacija) u limfni čvor
www.belimantil.info
155.
Migracija naivnih iefektornih T-limfocita
• Endotelne ćelije malih krvnih
sudova na mestu infekcije
(inflamacije) ispoljavaju adhezivne
molekule (E- i P- selektin, ICAM-1,
VCAM-1) koji se vezuju za
odgovarajuće molekule na
aktivisanim ili efektorskim T-
limfocitima
• Ekspresija adhezivnih molekula na
endotelu je pod uticajem TNF-α i IL-
1, koje produkuju tkivni makrofagi u
toku nespecifičnog imunog
odgovora na mikroorganizme
prodrle u tkivo
• E- i P- selektin u interakciji sa
odgovarajućim ligandima
omogućuju početnu slabu adheziju
(kotrljanje) limfocita
• Interakcija LFA-1ICAM-1 i VLA-
4VCAM-1 omogućuje čvrstu vezu i
sledstvenu transmigraciju
• Transmigracija se odvija pod
uticajem hemokina koje produkuju
leukociti na mestu inflamacije i
endotelne ćelije ili hemokina koji su
vezani za endotelne ćelije.
www.belimantil.info
156.
• Efektorski T-limfocitimigriraju na mesta
infekcije koristeći receptore koji
prepoznaju ligande ispoljene na endotelu
pod uticajem citokina produkovanih u toku
nespecifičnog imuniteta na
mikroorganizme
• Naivni T-limfociti nemaju ispoljene
receptore za ove ligande i zbog toga ne
mogu da migriraju na mesto infekcije
• T-limfociti koji prepoznaju antigene
mikroorganizama se zadržavaju u tkivu
posredstvom adhezionih interakcija
(interakcija VLA-4 integrina sa
fibronektinom, komponentom venćelijskog
matriksa)
• Aktivisani, antigen-specifični T-limfociti
koji su zadržani na mestu infekcije
obavljaju efektorske funkcije
• Aktivisani T-limfociti koji nisu specifični
za antigene mikroorganizama napuštaju
mesto infekcije preko limfatičkih sudova i
vraćaju se u cirkulaciju
www.belimantil.info
157.
EFEKTORSKE FUNKCIJE CD4LIMFOCITA
TH1 subpopulacija efektorskih CD4+ T-limfocita aktivira
makrofage koji su predhodno fagocitovali mikroorganizme
Aktivacija makrofaga je posredovana citokinima i direktnim
kontaktima
Aktivisani makrofagi povećavaju mikrobicidnu aktivnost i
sposobnost ubijanja fagocitovanih mikroorganizama
Idenični mehanizmi i reakcije se odigravaju nakon ubrizgavanja
mikrobskih proteina u kožu individue koja je predhodno
imunizovana tim mikrobom (prethodna infekcija ili vakcinacija)
www.belimantil.info
158.
Morfološke karakteristike
reakcije kasnepreosetljivosti
• Nakon intradermalnog ubrizgavanja
antigena sa kojim je organizam ranije
bio u kontaktu javlja se reakcija kasne
preosetljivosti (RKP)
• RKP se manifestuje 24-48 časova
nakon ubrizgavanja antigena, a
karakteriše se otokom, crvenilom i
otvrdnućem (induracijom)
• U RKP najčešće se primenjuje PPD
(purifikovani protein iz Mycobacteria)
radi procene specifičnog imuniteta
prema tuberkulozi
• Histološka karakteristika RKP je
prisusvo perivaskularnih ćelijskih
infiltrata sastavljenih od limfocita,
monocita i makrofaga, edem i
nakupljanje fibrina
• Ove patohistoloke promene su
posledica migracije CD4+ T-limfocita u
tkivo, migracije monocita u tkivo i
njihove transformacije u makrofage
kao i oštećenja tkiva produktima
aktivisanih makrofaga
www.belimantil.info
159.
AKTIVACIJA MAKROFAGA POMOĆUT-LIMFOCITA
Ćelijski imunitet je od ključnog značaja za eliminaciju
intracelularnih mikroorganizama kada:
sami makrofagi nisu aktivirani mikroorganizmima (neefikasan
urođeni imunitet)
patogeni mikroorganizmi izbegavaju odbrambene mehanizme
urođenog imuniteta
Dodatna aktivacija makrofaga pomoću T-limfocita je praćena:
eliminacijom intracelularnih mikroorganizama i brzom
rezolucijom infekcije
ili pod izvesnim uslovima
hroničnom inflamacijom po tipu RKP i formiranjem granuloma
kao i oštećenjem tkiva izazvanog toksičnim produktima makrofaga
Granulom čini kolekciju aktiviranih limfocita i makrofaga u
nekrotičnom tkivu koje kasnije fibrozira zbog proliferacije
fibroblasta i produkcije kolagena
www.belimantil.info
160.
Povećana
ekspresija MHC
molekula i
kostimulatora(B7
molekula)
Tkivni makrofagi fagocituju mikroorganizme. U fagolizozomima dolazi do prerade
proteinskih antigena do peptida koji se zatim ispoljavaju na membrani makrofaga u
kompleksu sa MHC molekulima II klase
Efektorski T-limfociti (koji u kontaktu sa makrofagom postaju TH1 ćelije) prepoznaju
antigenske peptide i bivaju aktivirani
Aktivirani TH1 limfociti ispoljavaju CD40L (CD154) i sekretuju citokine (IFN-γ)
Vezivanjem CD40L za CD40 na membrani makrofaga omogućava se aktivacija
makrofaga u bliskom kontaktu sa efektorskim TH1 limfocitima.
www.belimantil.info
161.
Povećana
ekspresija MHC
molekula i
kostimulatora(B7
molekula)
Vezivanjem IFN-γ za IFN-γ receptor na makrofagu dolazi do amplifikacije aktivacijskog
signala indukovanog posredstvom CD40-CD40L
Oba aktivaciona signala dovode do sinteze transkripcionih faktora koji aktiviraju gene za
lizozomalne proteaze, NO-sintazu i enzime koji sintetišu kiseoničke radikale (ROI)
Ovi enzimi dovode do ubijanja fagocitovanih mikroorganizama
Aktivirani makrofagi sekretuju citokine (TNF- α, IL-1 i hemokine) koji indukuju inflamaciju
i citokine koji aktiviraju T-limfocite (IL-12)
IL-12 povećava ekspresiju MHC molekula II klase i kostimulatora (B7 molekuli) čime se
obezbeđuje dodatna stimulacija T-ćelijskog imunog odgovorawww.belimantil.info
162.
OSTALE FUNKCIJE AKTIVISANIHEFEKTORSKIH CD4+ T-
LIMFOCITA
Produkcija TNF-α koji stimuliše:
- produkciju hemokina
- ekspresiju adhezivnih molekula na endotelu
Dodatna mobilizacija T-limfocita i leukocita (neutrofili i monociti) na
mesto infekcije i pospešivanje eradikacije infekcije
Pomoć CD8+ T-limfocitima za diferencijaciju u aktivirane citolitičke T-
limfocite
Pomoć B-limfocitima za diferencijaciju u plazmocite
www.belimantil.info
163.
FUNKCIJA CD8+ T-LIMFOCITAU AKTIVACIJI MAKROFAGA
Makrofage mogu aktivisati i CD8+ T-limfociti sličnim mehanizmima
kao i CD4+ T-limfociti.
CD8+ T-limfociti prepoznaju na makrofagama antigenske peptide u
sklopu MHC molekula I klase
- peptide produkovane od proteina iz citoplazme
- peptide koji potiču od fagocitovanih mikroba (mehanizam
ukrštene prezentacije)
Aktivacija makrofaga se odvija aktivacijom CD40-CD40L i IFN-γ-IFN
-γ receptor interakcijama
www.belimantil.info
164.
Dvosmerna komunikacija između
T-limfocitai makrofaga je odličan
primer povezanosti urođenog i
stečenog imuniteta
Makrofagi koji su stimulisani
procesom fagocitoze
mikroorganizama produkuju IL-12
IL-12 stimuliše diferencijaciju
naivnih CD4+ T-limfocita i njihovu
diferencijaciju u TH1 subpopulaciju
efektorskih ćelija koje sekretuju
IFN-γ.
IFN-γ aktivira makrofage koji
zatim ubijaju fagocitovane
mikroorganizme
www.belimantil.info
165.
KAKVA JE ULOGATH2 LIMFOCITA U ĆELIJSKOM IMUNITETU?
TH2 limfociti produkuju IL-4, IL-5, IL-10, IL-13.
IL-4 stimuliše produkciju IgE, a IL-5 aktivira eozinofile, važne komponente u odbrani od
infekcije helmintima
IL4, IL-10, IL-13 inhibiraju aktivaciju makrofaga, čime se zaustavlja RKP i ograničava
oštećenje tkiva izazvano protektivnim imunim odgovorom posredovanim TH1 limfocitima
Balans između TH1 i TH2 limfocita može da odredi ishod infekcije. Prikazani su neki
karakteristični primeriwww.belimantil.info
166.
EFEKTORSKE FUNKCIJE CD8+T-LIMFOCITA
Efektorski CD8+ T-limfociti vrše citolizu ciljnih ćelija i zato se zovu
citotoksični (citolitički) T-limfociti (CTL)
Mete citolitičkih T-limfocita su sopstvene ćelije koje ispoljavaju strane
antigenske peptide u sklopu MHC molekula I klase ili alogene ćelije
(transplantacija)
Antigenski peptidi potiču od proteina iz citoplazme (npr. virusni
proteini) ili iz vezikula (fagocitovani mikroorganizmi, fagocitovane
apoptotične ćelije) transportom u citoplazmu (ukrštena prezentacija)
Za citolizu je neophodan direktan međućelijski kontakt
Citoliza je antigen specifična
Za diferencijaciju naivnih CD8+ T-limfocita u efektorske CD8+ T-
limfocite neophodna je pomoć od strane CD4+ T-limfocita i kostimulacija
Diferentovani CTL ne zahtevaju pomoć od strane CD4+ T-limfocita i
kostimulaciju i zato oni mogu ubijati inficirane ćelije u bilo kom tkivu
www.belimantil.info
167.
• Mehanizam citolize(ubijanja inficiranih
ćelija) od strane CD8+ CTL prolazi kroz
nekoliko faza
• Prva faza je faza prepoznavanja antigena
na inficiranim ćelijama i formiranje
konjugata CTL-inficirana ćelija
• U ovoj fazi CTL svojim TCR-om
prepoznaje peptid mikroorganizma u žlebu
MHC molekula I klase. Koreceptorski CD8
molekul se veže za α-3 domen α lanca MHC
molekula I klase čime se obezbeđuje bolja
adhezija i prenos signala aktivacije
• U formiranju čvršće veze između CTL i
ciljne ćelije učestvuju adhezioni molekuli
od kojih je najznačajnija interakcija između
CD2 (CTL) i LFA-3 (CD58) (ciljna ćelija), kao
i LFA-1 (CD11aCD18) (CTL) i ICAM-1 (CD54)
(ciljna ćelija)
• Druga faza je faza aktivacije CTL u kojoj
prenos signala aktivacije unutar CTL
dovodi do egzocitoze sadržaja granula CTL
na mestima kontakta sa ciljnim ćelijama
www.belimantil.info
168.
• U trećojfazi dolazi do egzocitoze granula iz
CTL, i oslobađanja citolitičkih supstanci
• Najznačajnije citolitičke supstance su perforin,
granzimi, serin-proteaze i citokini (TNF-α i
Limfotoksin)
• Perforini se uglavljuju u membranu ciljne
ćelije, a zatim polimerizuju pod uticajem visoke
koncentracije Ca2+ jona prisutnih u
ekstraćelijskom prostoru
• Polimerizovani perforin formira pore na
membrani ciljnih ćelija kroz koje ulaze granzimi
• U četvrtoj fazi (faza ubijanja ciljne ćelije) dolazi
do apoptoze (programirane ćelijske smrti) ciljne
ćelije
• Apoptozu indukuju granzimi koji aktiviraju
seriju enzima u ciljnoj ćeliji (kaspaze).
• Kaspaze dovode do fragmentacije komponenti
jedra (nukleinske kiseline i proteini) i
sledstvene smrti ćelije.
• Apoptotski signal se može preneti i preko
interakcije Fas (CD95) molekula (ciljna ćelija)
vezivanjem za Fas-L (CD95L) (CTL)
• U petoj fazi dolazi do odvajanja CTL od ciljne
ćelije i vezivanje CTL za neku drugu ciljnu ćeliju
www.belimantil.info
169.
U makrofagu kojije inficiran intracelularnom bakterijom, neke od
bakterija su sekvestrirane u vezikulama (fagozomi), a druge mogu da
pređu u citoplazmu izbegnuvši fagolizozomalni put.
CD4+ T-limfociti prepoznaju antigene koji potiču od mikroorganizama iz
vezikula i aktiviraju makrofag da ubije mikroorganizme u vezikulama
CD8+ T-limfociti prepoznaju antigene koji potiču od bakterija iz
citoplazme, a zatim ubijaju inficirani makrofag čime se eliminiše rezervoar
infekcije
Kako efektorski CD4+ CD8+
T-limfociti sarađuju u
eradikaciji intracelularnih
infekcija?
www.belimantil.info
170.
PODSETNIK
Ćelijski imunitet jedeo stečenog imuniteta koji omogućava eradikaciju infekcija
izazvanih intracelularnim mikroorganizmima. U imunim reakcijama ćelijskog imuniteta
učestvuju CD4+ i CD8+ efektorski T-limfociti. CD4+ T-limfociti aktiviraju makrofage koje
zatim ubijaju fagocitovane mikroorganizme koji inače preživljavaju u vezikulama
fagocita. CD8+ CTL ubijaju ćelije u kojima se mikroorganizmi nalaze u citoplazmi, čime se
eliminiše rezervoar infekcije.
Efektorski T-limfociti se stvaraju od aktiivsanih naivnih T-limfocita u perifernim
limfoidnim organima, pretežno u limfnim čvorovima koji dreniraju tkiva u koje su prodrli
mikroorganizmi. Efektorski T-limfociti zatim migriraju na mesta infekcije.
Migracija efektorskih T-limfocita je kontrolisana adhezionim molekulima, koji se na
njima indukuju nakon aktivacije. Ovim molekulima T-limfociti se vezuju za odgovarajuće
ligande koji se ispoljavaju na endotelnim ćelijama pod uticajem mikroorganizama i
citokina produkovanih u toku nespecifičnog imunog odgovora na mikroorganizme.
Migracija T-limfocita je nezavisna od antigena, međutim, limfociti koji prepoznaju
antigene mikroorganizama u tkivu se zadržavaju u tkivu.
Efektorske ćelije TH1 subpopulacije CD4+ T-limfocita prepoznaju antigene
mikroorganizama koji su fagocitovani od strane makrofaga. Ovi T-limfociti ispoljavaju
CD40L i sekretuju IFN-γ koji zajedno dovode do aktivacije makrofaga
www.belimantil.info
171.
PODSETNIK
Aktivisani makrofagi produkujusupstance kao što su reaktivni kiseonički radikali
(intermedijeri), NO i lizozomalni enzimi koji ubijaju mikroorganizme. Makrofagi takođe
produkuju citokine koji indukuju inflamaciju, kao i citokine koji pospešuju fibrozu i
oporavak (ozdravljenje) tkiva
Efektorski CD4+ T-limfociti TH2 subpopulacije stimulišu eozinofilnu inflamaciju i
inhibiraju aktivaciju makrofaga. Eozinofili su važni u odbrani organizma od parazita
(helminti). Balans između aktivacije TH1 i TH2 ćelija određuje ishod mnogih infekcija, u
kojima TH1 limfociti stimulišu, a TH2 limfociti inhibiraju odbranu prema intracelularnim
mikroorganizmima
CD8+ T-limfociti se diferenciraju u CTL koji ubijaju inficirane ćelije, pretežno indukujući
fragmentaciju DNK i apoptozu
CD4+ i CD8+ T-limfociti često funkcionišu zajedno u eradikaciji intracelularnih infekcija
Mnogi patogeni mikroorganizmi su razvili mehanizme kojima se brane od ćelijskog
imuniteta. Ti mehanizmi su: inhibicija fuzije fagolizozoma, sposobnost izlaska iz vezikula
(fagozoma) u citoplazmu, inhibicija povezivanja kompleksa peptida i MHC molekula I
klase, produkcija inhibitornih citokina ili produkcija solubilnih citokinskih receptora.
www.belimantil.info
172.
PITANJA ZA PROVERUZNANJA
1. Koji su tipovi imunskih reakcija posredovanih T-limfocitima za
eliminaciju mikroorganizama koji su sekvestrirani u
vezikulama fagocita i mikroorganizama koji preživljavaju u
citoplazmi inficiranih ćelija domaćina?
2. Zašto diferntovani efektorski T-limfociti, prethodno aktivirani
antigenom pretežno migiriraju u tkiva na mestima infekcije, a
ne u limfni čvor i kojim se to mehanizmima postiže?
3. Pomoću kojih mehanizama efektorski T-limfociti aktiviraju
makrofage i kakav je odgovor makrofaga kojim se postiže
ubijanje fagocitovanih mikroorganizama?
4. Kakve su uloge TH1 i TH2 ćelija u odbrani od intracelularnih
mikroorganizama i parazita (helminata)?
5. Na koji način citolitički CD8+ T-limfociti ubijaju ćelije inficirane
virusima?
www.belimantil.info
173.
PODSETNIK
Ćelijski imunitet jedeo stečenog imuniteta koji omogućava eradikaciju
infekcija izazvanih intracelularnim mikroorganizmima
Efektorski T-limfociti se stvaraju od aktiivsanih naivnih T-limfocita u perifernim limfoidnim
organima, pretežno u limfnim čvorovima koji dreniraju tkiva u koje su prodrli
mikroorganizmi. Efektorski T-limfociti zatim migriraju na mesta infekcije.
Migracija efektorskih T-limfocita je kontrolisana adhezionim molekulima, koji se na njima
ispoljavaju nakon aktivacije. Ovim molekulima T-limfociti se vezuju za odgovarajuće
ligande koji se ispoljavaju na endotelnim ćelijama pod uticajem mikroorganizama i
citokina produkovanih u toku nespecifičnog imunog odgovora na mikroorganizme.
Migracija T-limfocita je nezavisna od antigena, međutim, samo limfociti koji u tkivu
prepoznaju antigene mikroorganizama se zadržavaju u tom tkivu.
www.belimantil.info
174.
1. Efektorski CD4+T-limfociti (Th) aktiviraju makrofage da ubijaju fagocitovane
mikroorganizme koji inače preživljavaju u vezikulama fagocita:
-Th1 subpopulacija ispoljava CD40L i sekretuje IFN-γ koji zajedno dovode do
aktivacije makrofaga. Aktivisani makrofagi produkuju supstance kao što su
reaktivni kiseonički radikali (intermedijeri), NO i lizozomalni enzimi koji ubijaju
mikroorganizme. Makrofagi takođe produkuju citokine koji indukuju inflamaciju, kao i
citokine koji pospešuju fibrozu i oporavak (ozdravljenje) tkiva
- Th2 subpopulacija stimuliše eozinofilnu inflamaciju i inhibira aktivaciju
makrofaga. Eozinofili su važni u odbrani organizma od parazita (helminti). Balans
između aktivacije Th1 i Th2 ćelija određuje ishod mnogih infekcija, u kojima Th1
limfociti stimulišu, a Th2 limfociti inhibiraju odbranu prema intracelularnim
mikroorganizmima
2. Efektorski CD8+ T limfociti (CTL) ubijaju ćelije u kojima se mikroorganizmi nalaze
u citoplazmi. Ubijanje inficiranih ćelija se pretežno vrši fragmentacijom jedra i
apoptozom
3. CD4+ i CD8+ T-limfociti često funkcionišu zajedno u eradikaciji intracelularnih
infekcija
________________________________________________________________________________
Mnogi patogeni mikroorganizmi su razvili mehanizme kojima se brane od ćelijskog
imuniteta. Ti mehanizmi su: inhibicija fuzije fagolizozoma, sposobnost izlaska iz
vezikula (fagozoma) u citoplazmu, inhibicija povezivanja kompleksa peptida i MHC
molekula I klase, produkcija inhibitornih citokina ili produkcija solubilnih
citokinskih receptora.
www.belimantil.info
175.
BCR KOMPLEKS
•Molekul antitela(imunoglobulina)
prepoznavanje antigena
•Nekovalentno vezani Igα i Igβ
prenos signala aktivacije
www.belimantil.info
176.
STRUKTURA ANTITELA
-At jesastavljeno od dva identična
teška (H) lanca i dva identična laka
(L) lanca
-V regioni i C regioni; regioni
komplementarnosti
-Fab fragment; Fc fragment
-Paratop/epitop
-Prepoznavanje proteina i
neproteina, nativnog molekula,
solubilnog i nesolubilnog antigena,
makromolekula ili malog molekula
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanjawww.belimantil.info
177.
HUMORALNI IMUNI ODGOVOR
Humoralniimuni odgovor je komponenta specifičnog imuniteta
koja je posredovana antitelima
Osnovne funkcije: neutralizacija i eliminacija mikroorganizama i njihovih
toksina
Antitela produkuju B-limfociti i njihovi progenitori
Nakon aktivacije B limfociti se difereciraju u efektorske ćelije,
plazmocite, koje sekretuju antitela.
www.belimantil.info
178.
Faza prepoznavanja: interakcijamembranskih imunoglobulina (IgM i IgD) iste specifičnosti na
naivnim B limfocitima sa antigenom.
Aktivacija naivnih B-limfocita pomoću antigena, Th limfocita i drugih dodatnih signala.
Proliferacija B-limfocita (klonska ekspanzija).
Diferencijacija u efektorske ćelije (plazmocite) koje aktivno sekretuju antitela; sekretovana
antitela su iste specifičnosti za antigen kao i membranski imunoglobulini na naivnom B-limfocitu.
U toku diferencijacije, osim produkcije IgM, neki B-limfociti mogu da sekretuju antitela drugih klasa
(izotipsko prekopčavanje). Ponovno izlaganje proteinskog antigena B-limfocitu dovodi do
produkcije antitela čiji je afinitet vezivanja za antigen povećan (afinitetno sazrevanje) Neki od B-
limfocita koji sekretuju visokoafintetna antitela diferentuju se u dugoživeće B-limfocite pamćenja.
Shema preuzeta iz knjige
Basic Immunology, Abbas i
Lichtman, iskljućivo za
potrebe predavanja
www.belimantil.info
179.
TIPOVI HUMORALNOG IMUNOGODGOVORA U ZAVISNOSTI OD
VRSTE ANTIGENA
T-zavisni imuni odgovor: odgovor na proteinske antigene (zavisan od pomoći
CD4+ T limfocita
T-nezavisni imuni odgovor: odgovor na neproteinske antigene (nije potrebna
pomoć CD4+ T limfocita)
www.belimantil.info
180.
Primarni humoralni imuniodgovor
nastaje pri prvom kontaktu sa antigenom
(primoinfekcija ili prva vakcinacija).
-U toku primarnog odgovora naivni
B-limfociti u perifernim limfoidnim organima se
aktiviraju, proliferišu i diferentuju u plazmocite i
memorijske B-limfocite.
-Neki plazmociti migriraju u kostnu
srž u kojoj preživljavaju duži period.
-Od aktivacije B limfocita antigenom
do razvoja primarnog humoralnog imunog
odgovora obično prođe 5-10 dana.
-Odgovor je relativno slab i praćen
sintezom antitela pretežno IgM klase a znatno
manje IgG klase.
-Afinitet vezivanja antitela za
antigen je relativno nizak.
Sekundarni humoralni imuni
odgovor nastaje pri ponovnom kontaktu sa
istim antigenom (reinfekcija, reimunizacija).
-Sekundarni humoralni imuni
odgovor karakteriše aktivacija memorijskih B
limfocita koji produkuju veće količine antitela.
On se razvija već posle 1-3 dana od aktivacije B
limfocita i karakterišu ga izraženiji procesi
izotipskog prekopčavanja i afinitetnog
sazrevanja (proteinski antigeni).
Sekretovana antitela su najčešće IgG klase ali
pod izvesnim uslovima mogu se sekretovati i
antitela IgA ili IgE klase.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
iskljućivo za potrebe predavanja
www.belimantil.info
181.
Vezivanje antigena zamembranske Ig na B-limfocitu
dovodi do grupisanja molekula BCR. Pri tome je
neophodno da se dva ili više receptora premoste
antigenom.
- Sledi prenos aktivacionih signala unutar B-limfocita
preko Ig-α i Ig-β (fosforilacija ITAM sekvenci pomoću
Fyn, Lyn i Blk tirozin kinaze.
- Za fosfotirozine na ITAM sekvencama priljubljuje se
Syk tirozin kinaza koja se takođe fosforiliše, što
omogućava vezivanje adaptornih proteina i njihovu
fosforilaciju. U narednom procesu stvaraju se brojni
biohemijski intermedijeri.
- Syk tirozin kinaza aktiviše PLC γ-1, koja preko
razgradnje membranskih inozitolfosfolipida (na ITP i
DAG) dovodi do aktivacije kalcijum-kalcineurinskog i
PKC aktivacionog puta.
-Signalni putevi koji započinju aktivacijom adaptornih
proteina stimulišu RAS-MAP kinazni put i Rac-JNK
aktivacioni put.
Dalji signalni proces preko aktivacije različitih
enzima je sličan aktivaciji T limfocita a njihova
posledica je sinteza i aktivacija različitih
transkripcionih faktora kao što su NFAT, Myc, NF-kB
i AP-1.
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, iskljućivo za potrebe predavanja
AKTIVACIJA
www.belimantil.info
182.
Kada je sistemkomplementa aktiviran
mikroorganizmima (ili polisaharidnim
antigenima) stvaraju se brojni fragmenti
komponenti komplementa.
Jedan od produkata, C3d, se vezuje za
receptor za C3d na B limfocitu (CR2,
CD21).
CR2 je tesno povezan sa CD19 i CD81
molekulima na B-limfocitima i oni zajedno
čine koreceptore na B ćelijama
Kada B-limfocit istovremeno prepozna
antigen mikroorganizma preko Ig
receptora i C3d vezanog za
mikroorganizam preko CR2 obezbeđena
su oba aktivaciona signala:
- Signal 1 se prenosi preko Ig-α i Ig-β
- Signal 2 se prenosi preko CD19
Sadejstvom oba aktivaciona signala
dolazi do aktivacije, proliferacije i
diferencijacije B-limfocita.
AKTIVACIJA
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanjawww.belimantil.info
183.
Aktivisani naivni B-limfocitiulaze u ćelijski
ciklus (proliferacija i formiranje klonova
limfocita)
Kada su u pitanju multivalentni
neproteinski antigeni, koji su takođe
potentni aktivatori komplementa. veći broj
B-limfocita se diferntuje u plazmocite koji
sekretuju IgM antitela.
Pošto su proteinski antigeni slabi
aktivatori B-limfocita imunski odgovor na
njih je slab. Međutim, oni indukuju promene
u B-limfocitima koje im omogućuju
interakciju sa T limfocitima:
- indukcija ekspresije B7 molekula
- indukcija receptora za citokine
- smanjenje ekspresije receptora za
hemokine što im onemogućava dalje
zadržavanje u folikulima i obezbeđuje
migraciju izvan folikula
PROLIFERACIJA / DIFERENCIJACIJA
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
184.
HUMORALNI IMUNI ODGOVORNA PROTEINSKE ANTIGENE
Humoralni imuni odgovor na proteinske antigene je zavisan od T-limfocita
pomoćnika (CD4+ Th limfocita)
U ovim procesima neophodan je kontakt između B limfocita i CD4+ T-limfocita
Ovaj tip imunskog odgovora se odvija u više faza:
1. Aktivacija T-limfocita proteinskim antigenom i diferencijacija T-limfocita u
efektorske ćelije
2. Aktivacija B-limfocita antigenom i prezentacija antigenskih peptida
efektorskim T-limfocitima
3. Aktivacija B-limfocita od strane efektorskih CD4+ T-limfocita
4. Diferencijacija aktivisanih B-limfocita u efektorske ćelije
5. Indukcija procesa izotipskog prekopčavanja
6. Indukcija procesa afinitetnog sazrevanja
www.belimantil.info
185.
1. CD4+ T-limfocitiprepoznaju u regionu parakorteksa limfnog čvora prerađene antigenske
peptide ispoljene na profesionalnim APC, nakon čega nastaje proliferacija i diferencijacija CD4+
T limfocita u efektorske CD4+ T-limfocite (Th ćelije). Th limfociti migriraju iz parakorteksa ka
limfnom folikulu.
2. Naivni B-limfociti, koji su u folikulu aktivisani antigenom migriraju do periferije folikula gde
dolaze u kontakt sa Th limfocitima- započinje T-B kooperacija.
Shema preuzeta iz
knjige Basic
Immunology, Abbas i
Lichtman, iskljućivo za
potrebe predavanja
www.belimantil.info
186.
B-limfociti prepoznaju nativniantigen u
folikulima i aktiviraju se:
-internalizacija antigena posredstvom BcR
-proteoliza po tipu prerade egzogenih
antigena
-ispoljavanje peptidnih fragmenata u
sklopu MHC molekula II klase
-ekspresija B7 molekula
CD4+ T-limfociti prepoznaju antigenske peptide u
sklopu MHC molekula II klase na B-limfocitima
- Signal 1 aktivacije: TcR/Ag peptid MHCII
- Signal 2 aktivacije: B7/CD28
nastanak efektorskih Th limfocita koji
ispoljavaju Cd40L i sekretuju citokine
__________________________________________
Ovim procesima omogućava se interakcija
limfocita koji su iste specifičnosti za antigen.
B-limfocit prepoznaje nativne (konformacione)
epitope na protienskom antigenu, dok CD4+ T-
limfocit prepoznaje peptidne fragmente tog istog
proteina.
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
187.
Vezivanjem CD40L odstrane CD40 molekula na B-limfocitu kao i vezivanjem citokina preko
receptora za citokine na B-limfocitu omogućena je proliferacija (klonska ekspanzija) B-limfocita i
njihova diferencijacija u efektorske ćelije koje sekretuju antitela.
Izotipsko prekopčavanje (“switch”): stvaranje antitela različitih izotipova (klasa)
Afinitetno sazrevanje: stvaranje antitela većeg afiniteta za antigen
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
188.
U B-limfocitu kojisekretuje antitela
IgM klase došlo je do preuređenja
genskih segmenata (VDJ povezan sa
najbližim Cμ genskim segmentom;
preuređnje VDJ se dešava na nivou
DNK a povezivanje VDJ sa Cμ na
nivou primarnog transkripta
Signali od strane T-limfocita
(ligacija CD40, citokini) indukuju
rekombinaciju prekopčavajućih
(“switch”) regiona ; switch (S)
regioni(Sμ, Sγ, Sε, Sα) su
konzervisane sekvence nukleotida
smeštene ispred odgovarajućih C
gena. Jedino Cδ nema S sekvenceU
toku procesa prekopčavanja (na
nivou primarnog transkripta) Sμ se
povezuje sa odgovarajućim S
sekvencama drugih C gena.Svi ostali
nukleotidi između S sekvenci se
uklanjaju procesom delecije
________________________________
Prikazan je primer izotipskog
prekopčavanja za sintezu teškog lanca IgG i
teškog lanca IgE
Izotipsko prekopčavanje
(“switch”): produkcija antitela
različitih klasa
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo
za potrebe predavanjawww.belimantil.info
189.
Za sekreciju IgM,čija je osnovna efektorska funkcija aktivacija komplementa, nije neophodna pomoć od
strane T-limfocita.
Pod uticajem CD40L ispoljenog na pomoćničkim T-limfocitima i citokina koje ove ćelije produkuju
dolazi do procesa izotipskog prekopčavanja u B-limfocitima i sekrecije drugih klasa antitela.
Na produkciju i sekreciju IgG (podklase IgG 1 i IgG3) utiče IFN-γ; ova antitela učestvuju u procesima
opsonizacije i fagocitoze (posredovani Fc receptorom), u aktivaciji komplementa i neonatalnom
imunitetu.
Na produkciju i sekreciju IgE koji je bitan za imunitet prema helmintima i reakciji ranog tipa
preosetljivosti utiče Il-4.
Na produkciju i sekreciju IgA u mukoznim tkivima koji su važni za lokalni imunitet sluznica utiče TGF β.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
190.
Povećanje afiniteta antitelau toku humoralnog
imunog odgovora na proteinski antigen, koje se
javlja pri produženom ili ponovljenom izlaganju
tog antigena imunom sistemu :
- Posledica je tačkastih mutacija u
hipervarijabilnim regionima V gena Ig
- Ove mutacije se zovu somatske
hipermutacije zbog toga što su i do 1000 puta
češće nego tačkaste mutacije u mnogim drugim
genima (1 mutacija na 103 baznih parova po
deobi)
Dešavaju se u B-limfocitima u germinativnim
centrima folikula perifernih limfoidnih organa i
tkiva
Neki B-limfociti aktivisani
proteinskim antigenom i Th-limfocitima
migriraju u folikul gde ubrzano proliferišu i
formiraju germinativni centar; zbog somatskih
hipermutacija ovi B-limfociti produkuju antitela
(membranski Ig) različitog afiniteta vezivanja za
antigen.
FDC vezuju antigene u obliku imunskih
kompleksa, a takođe i C3b i C3d komponentu
komplementa preko CR2
Jedino B-limfociti sa visokim afinitetom za
antigen mogu da se vežu za FDC i da od FDC
(preko CD40-L) prime signal za preživljavanje
Ćelije sa nisko afinitetnim receptorom za
antigen umiru procesom apoptoze
Afinitetno sazrevanje
www.belimantil.info
191.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe
predavanjawww.belimantil.info
192.
1. U inicijalnojfazi primarnog humoralnog imunog odgovora aktivacija B-limfocita se dešava u folikulima,
a T-limfocita u T-zavisnim zonama.
2. Aktivirani T- i B-limfociti migriraju i susreću se na granici folikula i T-zavisnih zona.
3. U toku T-B koooperacije dolazi do proliferacije, izotipskog prekopčavanja i diferencijacije B-limfocita u
plazmocite. Plazmociti ostaju u perifernim limfoidnim organima izvan folikula, a jedan broj migrira u
kostnu srž (dugoživeći plazmociti).
4. Neki aktivisani B-limfociti se vraćaju u folikul (germinativne centre).
5. U germinativnim centrima dolazi do afinitetnog sazrevanja i dodatnih procesa izotipskog
prekopčavanja kao i formiranja dugoživećih B-limofcita pamćenja. B-limfociti pamćenja poseduju
preuređene gene za teške lance Ig i ispoljene visokoafinitetne receptore za antigen. Sami ne produkuju
antitela.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe
predavanja
www.belimantil.info
193.
-Sekretovana IgG antitela
formirajuimune komplekse sa
preostalim antigenom.
-Kompleksi reaguju sa B-
limfocitima preko BCR-a i preko
receptora za Fc fragmenta antitela,
FcγRIIB (CD32).
-Za CD32 su vezane
fosfataze koje uklanjaju fosfatne
grupe sa ITAM motiva Igα i Igβ
molekula i na taj način se blokira
prenos signala preko BCR
REGULACIJA HUMORALNOG
IMUNOG ODGOVORA-
zaustavljanje humoralnog imunog
odgovora:
- apoptoza plazmocita
- mehanizam povratne sprege
posredstvom antitela
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
194.
EFEKTORSKI MEHANIZMI HUMORALNOG
IMUNITETA
Zaštitaod ekstracelularnih mikroorganizama i njihovih toksina: vezivanjem za
mikroorganizme i toksine sprečava se infekcija i oštećenje ćelija; antitela su jedina
komponenta specifičnog imuniteta koja sprečava infekciju
Antitela pomažu u eliminaciji potencijalno štetnih mikroorganizama i agenasa
Antitela mogu da obavljaju svoje efektorske funkcije udaljeno od mesta gde se
stvaraju
Najveći broj vakcina se zasniva na indukciji specifičnog humoralnog imuniteta
Posledice defekta humoralnog imuniteta su česte infekcije izazvane bakterijama,
virusima i parazitima
www.belimantil.info
195.
KARAKTERISTIKE ANTITELA KOJEODREĐUJU NJIHOVE
EFEKTORSKE FUNKCIJE
Fab fragmentom antitela vezuju i blokiraju štetne efekte mikroorganizama i njihovih
toksina
Fc fragmentom antitela aktiviraju različite efektorske mehanizme kojima se eliminišu
mikroorganizmi i njihovi toksini
__________________________________________________________________________
Protektivna antitela se produkuju u toku primarnog imunog odgovora, a u većoj
koncentraciji u toku sekundarnog imunog odgovora; mehanizmi izotipskog
prekopčavanja i afinitetnog sazrevanja povećavaju protektivne funkcije antitela
EFEKTORSKE FUNKCIJE ANTITELA:
1. Neutralizacija mikroorganizama i toksina
2. Opsonizacija antigena za fagocitozu od strane makrofaga i neutrofila
3. Aktivacija klasičnog puta komplementa
4. Citotoksičnost zavisna od antitela (ADCC) posredovana NK ćelijama i eozinofilima
5. Reakcija ranog tipa preosetljivosti
6- Imunitet novorođnečeta
7. Mukozni imunitet
8. Regulacija humoralnog imunog odgovora
www.belimantil.info
196.
EFEKTORSKE FUNKCIJE ANTITELASPECIFIČNE ZA IZOTIP:
IgG: - neutralizacija mikroba i toksina
- opsonizacija za fagocitozu
- aktivacija klasičnog puta komplementa
- ADCC od strane NK ćelija
- neonatalni imunitet
- regulacija humoralnog imunog odgovora
IgM: - aktivacija klasičnog puta komplementa
IgA: - mukozni imunitet
- neutralizacija mikroba i toksina
IgE: - ADCC od strane eozinofila
- rani tip preosetljivosti
www.belimantil.info
197.
A. Antitela sprečavaju
vezivanjemikroba za
ćelije. Na taj način se
sprečava
sposobnost mikroba
da inficira ćelije
domaćina
B. Antitela inhibiraju
širenje infekcije sa
inficirane ćelije na
drugu, inficiranu
ćeliju
C. Antitela sprečavaju
vezivanje toksina za
ćeliju i tako
inhibiraju patogene
efekte toksina.
Fab funkcija
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanjawww.belimantil.info
198.
Opsonizacija – oblaganje
antigena(mikroorganizma)
antitelom (Fab) ili nekim
drugim molekulom. Opsonini –
molekuli koji vrše opsonizaciju
(najčešće su antitela IgG
klase); opsonizacija je priprema
za fagocitozu.
Fagociti ispoljavaju receptor za
Fc fragment opsonizujućeg
antitela CD64 (FcγRI);
angažovanjem FcγRI aktiviraju
se fagociti nakon čega dolazi
do fagocitoze i ubijanja
fagocitovanih mikroba.
___________________________
Značajan mehanizam odbrane
od enkapsuliranih bakterija
(pneumokok) koji imaju
polisaharidni zid.
Fab/Fc funkcija
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanja
www.belimantil.info
199.
Neke potklase IgGvezuju
svojim Fab fragmentom
antigene na ćelijama (npr.
inficirane ćelije).
NK ćelije ispoljavaju CD16
(FcγRIII) kojim vežu Fc
fragment antitela;
angažovanjem CD16 aktivira se
NK ćelija koja ubija ciljnu ćeliju
IgE se vežu Fab fragmentom za
antigene na površini parazita
(helminata).
Eozinofili ispoljavaju
visokoafinitetne receptore za
Fc fragment IgE (FcεRI);
angažovanjem FCεRI dolazi do
aktivacije eozinofila koje
oslobađaju granule (toksični
medijatori granula ubijaju
parazite).
Fab/Fc funkcija
(ADCC)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
200.
- U sluzokožigastrointestinalnog i respiratornog trakta plazmociti (lamina propria)
produkuju i sekretuju IgA i vezni (J) lanac koji služi za formiranje dimernih molekula IgA
- Epitelne ćelije mukoza sekretuju poli-Ig receptor koji veže IgA (može i IgM)
- Kompleks IgA poli-Ig receptor se transportuje kroz epitelnu ćeliju; na površini epitelne
ćelije dolazi do delimične proteolize poli-Ig receptora i transporta IgA dimera i dela poli-Ig
receptora u lumen
____________________________________________________________________________________
Mukozni imunitet je efikasan u zaštiti od poliomijelitisa (oralni put unošenja vakcine).
(mukozni imunitet)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanja
Fab funkcija
www.belimantil.info
201.
Antitela majke (IgG)se aktivno transportuju kroz placentu do fetusa; ćelije
placente sekretuju neonatalni Fc receptor (FcRn) i kroz epitel digestivnog trakta
novorođenčeta
Antitela majke (IgG) iz mleka se aktivno transportuju kroz epitel digestivnog trakta
novorođenčeta; ćelije epitela digestivnog trakta novorođenčeta ispoljavaju Fc
receptor kojim vezuju antitela iz majčinog mleka.
IMUNITET NOVOROĐENČETA
-zaštita od infekcije (pasivni prirodni imunitet)-
www.belimantil.info
202.
Komplement (sistem komplementa)čini skup cirkulišućih i membranskih proteina
koji imaju važnu ulogu u odbrani organizma od mikroba i oštećenju tkiva
posredovanim antitelima.
Aktivacija sistema komplementa bez učešća antitela (ALTERNATIVNI I LEKTINSKI
PUT) je važna komponenta urođenog imuniteta prema infekciji.
Aktivacija komplementa uz učešće antitela (KLASIČNI PUT) je važan efektorski
mehanizam humoralnog imuniteta.
Aktivacija sistema komplementa: kaskada proteolitičke razgradnje proteina i
njihova amplifikacija.
Nekontrolisanu aktivaciju i sprečavanje oštećenja sopstvenih ćelija vrše regulatorni
proteini komplementa.
AKTIVACIJA SISTEMA KOMPLEMENTA
www.belimantil.info
203.
ALTERNATIVNI PUT aktivacije
-spontanmproteolizom C3, nastaje C3a i
C3b; C3b se vezuje za površinu mikroba
- za kompleks mikrob-C3b se vezuje
faktor B
- nastaje proteoliza faktora B i formiranje
kompleksa mikrob-C3bBb ( C3
konvertaza alternativog puta )
-dejstvom C3 konvertaze se dodatno
cepa C3, C3b se dodatno vezuje za
mikrob i formira se kompleks C3bBbC3b
(C5 konvertaza aletrnativnog puta)
KLASIČNI PUT aktivacije
- vezivanje C1 komponente za IgG
vezanog za površinu mikroba.
-proteoliza C4 i C2 komponenti i
formiranje komponente C4b2a
vezane za antitelo (C3 konvertaza
klasičnog puta)
- proteoliza C3 komponente i
formiranje C4b2aC3b (C5
konvertaza klasičnog puta)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za
potrebe predavanjawww.belimantil.info
204.
Kasna faza aktivacije(zajednička
za sve puteve aktivacije
komplementa) započinje
aktivacijom C5 konvertaze:
-Cepanje C5 komponente na C5a i
C5b
- C5b započinje formiranje
membranskog atakujućeg
kompleksa (MAC) vezivanjem za
C6 komponentu za koju se zatim
vezuje C7 komponenta
- C7 omogućuje ugradnju MAC
kompleksa u lipidnu membranu i
vezivanje C8 komponente.
- C8 omogućuje vezivanje C9 i
njenu polimerizaciju.
- Polimerizacijom C9 formira se
funkcionalni MAC kompleks (C5b,
C6, C7, C8, polimerizovani C9)
koji vrši perforaciju membrane
ćelije i sledstvenu lizu ćelije.
Shema preuzeta iz knjige Basic Immunology,
Abbas i Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
205.
Opsonizacija i fagocitoza
Komplementom
posredovanacitoliza
Stimulacija zapaljenske
reakcije.
Aktivacija humoralnog
imuniteta:
-Interakcija C3d sa
CR2 receptororom
na B-limfocitu.
-Selekcija
visokoafinitetnih B-
limfocita (vezani
proteinikomplementa
u sklopu imunskih
kompleksa bivaju
perpoznati od CR
ispoljenih na FDC).
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe predavanjawww.belimantil.info
206.
DAF (faktor ubrzanjarazgradnje): sprečava vezivanje faktora B za C3b ili vezivanje C4b2a za
C3b (zaustavljanje aktivacije alternativnog i klasičnog puta).
MCP (membranski kofaktorski protein): služi kao kofaktor sa faktorom I za proteolizu C3b na
inaktivne fragmente.
CR I (komplementni receptor tip I): vrši obe funkcije koje imaju DAF i MCP.
C1 INH (C1 inhibitor): zaustavlja ranu aktivaciju klasičnog puta na nivou C1.
Ostali regulatori (regulatori kasne faze aktivacije)
HEREDITARNI
ANGIONEUROTSKI EDEM:
deficijencija C1 INH
PAROKSIZMALNA NOĆNA
HEMOGLOBINURIJA: defekt
u sintezi brojnih glikolipida
uključujući i DAF i MCP
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, iskljućivo za potrebe
predavanja
www.belimantil.info
207.
-Sekretovana IgG antitela
formirajuimune komplekse sa
preostalim antigenom.
-Kompleksi reaguju sa B-
limfocitima preko BCR-a i preko
receptora za Fc fragmenta antitela,
FcγRIIB (CD32).
-Za CD32 su vezane
fosfataze koje uklanjaju fosfatne
grupe sa ITAM motiva Igα i Igβ
molekula i na taj način se blokira
prenos signala preko BCR
REGULACIJA HUMORALNOG
IMUNOG ODGOVORA-
zaustavljanje humoralnog imunog
odgovora:
- apoptoza plazmocita
- mehanizam povratne sprege
posredstvom antitela
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, iskljućivo za potrebe predavanja
www.belimantil.info
208.
PODSETNIK
1. Fiziološka ulogaimunog sistema je da uklanja tumorske ćelije i spreči rast
tumora.
2. Tumorski antigeni mogu biti proizvod onkogena ili tumor-supresorskih gena,
mutiranih ćelijskih proteina, preterano ili aberantno ispoljenih molekula, ili
proizvodi onkogenih virusa.
3. Odbacivanje tumora se vrši posredstvom CTL: Indukcija CTL odgovora
protiv tumorskih antigena često uključuje ingestiju tumorskih
ćelija/antigena i prezentaciju od strane profesionalnih APC.
4. Tumori izbegavaju imuni odgovor: prestaju da ispoljavaju tumorske
antigene, MHC molekule ili molekule uključene u preradu antigena,
sekretuju supresorske citokine.
5. Imunoterapija tumora: pasivna imunizacija, aktivna imunizacija (vakcinacija
tumorskim ćelijama/antigenima, vakcinacija genski izmenjenim tumorskim
ćelijama-kostimulatori, citokini, vakcinacija in vitro tretiranim DC)
www.belimantil.info
209.
6. Glavni uzrokodbacivanja transplantata su alogeni MHC (HLA)
antigeni.
7. T limfociti prepoznaju alogeni MHC (HLA) direktno (na APC davaoca)
ili indirektno (na APC primaoca-prerađen i u kompleksu sa
sopstvenim MHC)
8. Odbacivanje transplantata može biti: hiperakutno (preformirana
antitela), akutno (CTL) i hronično (CTL, Th limfociti)
9. Imunosupresija pri transplantaciji tkiva: antiaktivacioni,
antiproliferativni, antiinflamatorni lekovi.
10. Transplantacija kosne srži: mogućnost reakcije kalem-protiv-
domaćina.
www.belimantil.info
210.
REAKCIJE PREOSETLJIVOSTIREAKCIJE PREOSETLJIVOSTI
OŠTEĆENJATKIVA I POJAVA BOLESTI IZAZVANIH
REAKCIJAMA IMUNOG SISTEMA:
NEKONTROLISANI I POREMEĆENI IMUNI ODGOVOR NA STRANE
ANTIGENE
IMUNI ODGOVOR NA SOPSTVENE ANTIGENE
P O D E L A:
I TIP PREOSETLJIVOSTI: POSREDOVAN IgE ANTITELIMA I
MEDIJATORIMA IZ MASTOCITA
II TIP PREOSETLJIVOSTI: POSREDOVAN ANTITELIMA PREMA TKIVNIM
ANTIGENIMA
III TIP PREOSETLJIVOSTI: POSREDOVAN DEPONOVANJEM IMUNSKIH
KOMPLEKSA
IV TIP PREOSETLJIVOSTI: POSREDOVAN T LIMFOCITIMA
www.belimantil.info
211.
Tip preosetljivosti Patološkiimuni mehanizmi Mehanizmi oštećenja tkiva
TIP I: rana
preosetljivost
posredovana IgE
antitelima
TIP II:
preosetljivost
posredovana
antitelima
TIP III:
preosetljivost
posredovana
imunim
kompleksima
TIP IV:
preosetljivost
posredovana T
limfocitima
Shema preuzeta iz knjige Basic Immunology, Abbas
i Lichtman, isključivo za potrebe predavanja
Medijatori iz mastocita
(vazoaktivni amini, lipidni
medijatori, citokini)
-Angažovanje i aktivacija
nautrofila i makrofaga
posredstvom komplementa i
receptora za Fc
-Opsonizacija, fagocitoza
-Poremećaji funkcije
Angažovanje i aktivacija
leukocita posredstvom
komplementa i Fc receptora
-Aktivacija makrofaga,
inflamacija posredovana
citokinima
-- Direktna citoliza i
inflamacija posredstvom
citokina
www.belimantil.info
212.
I TIP PREOSETLJIVOSTIITIP PREOSETLJIVOSTI
RANA PREOSETLJIVOST, ALERGIJA
ALERGENI : antigeni iz okruženja koji pokreću alergijske reakcije
ATOPIJA : genetska sklonost ka alergijskim reakcijama
( predominacija Th2 odgovora)
ALERGIJSKE BOLESTI (relativno česte, oko 20% populacije)
POLENSKA KIJAVICA
BRONHIJALNA ASTMA
ALERGIJA NA HRANU
URTIKARIJA
ALERGIJA NA LEKOVE
ALERGIJA NA PRODUKTE UBODA INSEKATA
SISTEMSKA ANAFILAKSA
www.belimantil.info
213.
TOK REAKCIJE PREOSETLJIVOSTII TIPATOK REAKCIJE PREOSETLJIVOSTI I TIPA
1. SENZIBILIZACIJA
STIMULACIJA Th2 ODGOVORA NA ALERGEN
PRODUKCIJA IgE ANTITELA
VEZIVANJE IgE ANTITELA ZA MASTOCITE I BAZOFILE U KRVI
POSREDSTVOM VISOKOAFINITETNIH RECEPTORA IgE (Fc εRI)
2. PONOVNI KONTAKT SA ALERGENOM
PREMOŠĆAVANJE Fab FRAGMENTA IgE ANTITELA ALERGENOM
AKTIVACIJA MASTOCITA I BAZOFILA
OSLOBAĐANJE MEDIJATORA IZ MASTOCITA I BAZOFILA
3. EFEKAT MEDIJATORA
www.belimantil.info
214.
Prvo izlaganje antigenu
(alergenu)
AktivacijaTh2 ćelija i
stimulacija produkcije
(switch( IgE antitela od
strane B limfocita-
plazmocita
Vezivanje IgE za
visokoafinitetne FcεRI na
mast ćelijama i li bazofilima
Ponovljena izlaganja
alergenu
Aktivacija mastocita,
oslobađanje medijatora
Rana reakcija (minuti posle Kasna reakcija
ponovnog izlaganja alergenu) (6-24 sati posle ponovnog izlaganja)
Shema preuzeta iz knjige Basic Immunology,
Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
215.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanja
Mastociti senzibilisani
vezivanjem IgE za FcεRI
((A); Premošćavanje
receptora prilikom
ponovnog kontakta sa
alergenom i aktivacija
mastocita
DEGRANULACIJA
MASTOCITA:
oslobađanje medijatora
www.belimantil.info
Dilatacija Oštećenje DilatacijaKontrakcija Inflamacija
krvnih sudova tkiva krvnih sudova glatke muskulature (mobilizacija
leukocita
Kontrakcija
glatke muskulature
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
218.
T E RA P I J AT E R A P I J A
- ANTIHISTAMINICI
- EPINEFRIN (ADRENALIN)
- KORTIKOSTEROIDI
- HIPOSENZIBILIZACIJA (DESENZITIZACIJA)
(INDUKCIJA ALERGEN SPECIFIČNE TOLERANCIJE)
- ANTI – IgE ANTITELA
- INHIBITORI DEGRANULACIJE MASTOCITA (CROMOLIN)
www.belimantil.info
219.
II TIP PREOSETLJIVOSTIIITIP PREOSETLJIVOSTI
PREOSETLJIVOST IZAZVANA ANTITELIMA PREMA ĆELIJSKIM ILI TKIVNIM
ANTIGENIMA
ANTIGENI NA POVRŠINI ĆELIJA
KOMPONENTE VANĆELIJSKOG MATRIKSA
ANTIGENI:
- SOPSTEVNI MOLEKULI
- SOPSTVENI IZMENJENI MOLEKULI
- HAPTENI (LEKOVI)
MEHANIZAM OŠTEĆENJA:
- OPSONIZACIJA I FAGOCITOZA ĆELIJA
- INFLAMACIJA POSREDOVANA KOMPLEMENTOM I Fc RECEPTOROM
ANTITELA
- POREMEĆAJI FUNKCIJE ĆELIJA
www.belimantil.info
220.
BOLESTI IZAZVANE IITIPOM PREOSETLJIVSOTIBOLESTI IZAZVANE II TIPOM PREOSETLJIVSOTI
Bolest Ciljni antigen Mehanizam bolesti Kliničke manifestacije
Autoimuna hemolitička
anemija
Proteini membrane
eritrocita
Opsonizacija i fagocitoza
eritrocita
Hemoliza, anemija
Autoimuna
trombocitopenijska
purpura
Proteini membrane
trombocita
Opsonizacija i fagocitoza
eritrocita
Krvarenje
Pemfigus vulgaris Proteini na spojevima
epidermalnih ćelija
Aktivacija proteaza,
oštećenja međućelijskih veza
Kožne vezikule
(bule)
ANCA
Vaskulitis
Proteini granula
neutrofila
Degranulacija neutrofila i
inflamacija
Vaskulitis
Goodrasture-ov sindrom Nekolagneski proteini
glomerula bubrega i
alveola pluća
Inflamacija posredovana
komplementom i Fc
receptorom antitela
Nefritis,
plućna hemoragija
Akutna reumatska
groznica
Antigen ćelijskog zida
streptokoka (ukrštena
reaktivnost sa antigenom
srca)
Inflamacija
Aktivacija makrofaga
Miokarditis,
artritis
www.belimantil.info
III TIP PREOSETLJIVOSTIIIITIP PREOSETLJIVOSTI
OŠTEĆENJE TKIVA IZAZVANO IMUNSKIM KOMPLEKSIMA
- STVARANJE IMUNSKIH KOMPLEKSA U VIŠKU
- STVARANJE MALIH IMUNSKIH KOMPLEKSA
- NEEFIKASNO UKLANJANJE IMUNSKIH KOMPLEKSA
- DEPONOVANJE NA KARAKTERISTIČNIM MESTIMA
GLOMERULI BUBREGA
SINOVIJALNA MEMBRANA
KOŽA
POKRETANJE ZAPALJENSKE REAKACIJE
(VASCULITIS)
www.belimantil.info
223.
BOLESTIBOLESTI
- SERUMSKA BOLEST
-SISTEMSKI LUPUS ERYTHEMATOSUS
- POSTSTREPTOKOKNI GLOMERULONEFRITIS
- POLYARTHRITIS NODOSA
- ARTHUSOVA REAKCIJA (EKSPERIMENTALNI MODEL
VASCULITISA
MEHANIZMI OŠTEĆENJA TKIVA
INFLAMACIJA POSREDOVANA KOMPLEMENTOM I Fc
RECEPTOROM
www.belimantil.info
224.
Oštećenje izazvano anti-tkivnim
antitelima
Oštećenjetkiva izazvano imunim kompleksima
Mehanizmi deponovanja
antitela
Efektorski mehanizmi
oštećenja tkiva
Tipovi bolesti posredovanih antitelima
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
Tip II
preosetljivosti
Tip III
preosetljivosti
www.belimantil.info
225.
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
Inflamacija i oštećenje tkiva
(tip III)
Opsonizacija/fagocitoza
(tip II)
Interferencija (inhibicija ili
stimulacija) sa normalnom
funkcijom ćelija
(tip II)
www.belimantil.info
226.
IV TIP PREOSETLJIVOSTIIVTIP PREOSETLJIVOSTI
REAKCIJA KASNE PREOSETLJIVOSTI
OŠTEĆENJE TKIVA IZAZVANO T LIMFOCITIMA
CD4 (TH1) LIMFOCITI
CD8 LIMFOCITI
INDUKTORI: PROTEINSKI ANTIGENI I HEMIJSKE SUPSTANCE KOJE SE VEZUJU
ZA SOPSTVENE PROTEINE
STIMULATORI: CITOKINI Th1 LIMFOCITA
EFEKTORSKE ĆELIJE: CD8+ CITOTOKSIČNI LIMFOCITI
AKTIVISANI MAKROFAGI
MEHANIZAM OŠTEĆENJA TKIVA
AKUTNO: DELOVANJE PRODUKATA AKTIVISANIH MAKROFAGA
• PROTEOLITIČKI ENZIMI
• KISEONIČKI RADIKALI
• NO
• PROINFLAMATORNI CITOKINI
HRONIČNO: FIBROZA
• CITOKINI I FAKTORI RASTA PRODUKOVANI OD STRANE MAKROFAGA
www.belimantil.info
227.
Morfološke karakteristike
reakcije kasnepreosetljivosti
• Nakon intradermalnog ubrizgavanja
antigena sa kojim je organizam ranije
bio u kontaktu javlja se reakcija kasne
preosetljivosti (RKP)
• RKP se manifestuje 24-48 časova
nakon ubrizgavanja antigena, a
karakteriše se otokom, crvenilom i
otvrdnućem (induracijom)
• U RKP najčešće se primenjuje PPD
(purifikovani protein iz Mycobacteria)
radi procene specifičnog imuniteta
prema tuberkulozi
• Histološka karakteristika RKP je
prisusvo perivaskularnih ćelijskih
infiltrata sastavljenih od limfocita,
monocita i makrofaga, edem i
nakupljanje fibrina
• Ove patohistoloke promene su
posledica migracije CD4+ T-limfocita u
tkivo, migracije monocita u tkivo i
njihove transformacije u makrofage
kao i oštećenja tkiva produktima
aktivisanih makrofaga
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
isključivo za potrebe predavanjawww.belimantil.info
Shema preuzeta izknjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
A. Kasni tip (IV) preosetljivosti gde oštećenje tkiva nastaje aktivacijom CD4+ i CD8+ T ćelija koje
zatim aktiviraju makrofage i inflamatorne ćelije
B. Direktno ubijanje ciljnih ćelija posredstvom CD8+ CTL
www.belimantil.info
230.
•IMUNOLOŠKA TOLERANCIJA
•AUTOIMUNOST
Razlikovanje sopstvenogod stranog
i poremećaji diskriminacije
IMUNOLOŠKA TOLERANCIJA: izostanak imunog odgovora
(imunološka nereaktivnost) na neki (bilo koji) antigen
Autotolerancija: imunološka nereaktivnost na vlastite antigene
www.belimantil.info
231.
Kontakt antigena saatigen-specifičnim limfocitima može da ima tri ishoda:
1. Aktivacija limfocita i imuni odgovor (Imunogeni antigen)
2. Funkcionalna inaktivacija ili smrt limfocita: tolerancija (tolerogeni antigen); tolerancija na taj
antigen znači da će limfociti biti nereaktivni čak iako se naknadno unese isti antigen u imunogenoj
formi.
3. Ignorisanje: limfociti ignorišu prisutan antigen i ne reaguju (neimunogeni antigen); pri ponovnom
kontaktu limfociita sa tim antigenom u imunogenoj formi, može doći do imunog odgovora.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
232.
Da li ćenastati imuni odgovor (aktivacija), tolerancija ili
ignorisanje, određeno je:
-prirodom antigen-specifičnih limfocita
-prirodom antigena
-načinom prikazivanja antigena imunom sistemu
Isti antigen se može prikazivati na način i u obliku koji dovodi do
imunog odgovora ili tolerancije.
www.belimantil.info
233.
ZNAČAJ IMUNOLOŠKE TOLERANCIJE:
1.Autotolerancija: sopstveni antigeni normalno indukuju
toleranciju
2. Indukcija tolerancije limfocita na određeni antigen može
biti korisna u sprečavanju ili kontroli neželjene imunološke
reakcije:
- lečenje alergija
- lečenje autoimunih bolesti
- prevencija odbacivanja transplantiranih organa
www.belimantil.info
234.
AUTOTOLERANCIJA
Imunološka tolerancija na
sopstveneantigene može se
indukovati kada nezreli limfociti
prepoznaju sopstvene antigene u
primarnim limfoidnim organima:
centralna tolerancija, ili kada zreli
limfociti prepoznaju sopstvene
antigene u perifernim limfoidnim
tkivima: periferna tolerancija.
-centralna tolerancija je mehanizam
tolerancije prema sopstvenim
antigenima prisutnim u primarnim
limfoidnim organima (timus, kostna
srž).
- periferna tolerancija je mehanizam
tolerancije prema sopstvenim
antigenima koji nisu prisutni u
timusu ili kostnoj srži.
Shema preuzeta iz knjige Basic Immunology, Abbas
i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
235.
CENTRALNA TOLERANCIJA T
LIMFOCITA
CD4+Th limfociti su ključni za sve
vidove imunog odgovora
(humoralni/ćelijski) prema
proteinskim antigenima
Ako nezrele T ćelije prepoznaju
sopstvene antigene u timusu
visokim afinitetom, ove ćelije će
umreti apoptozom (negativna
selekcija).
Jaka interakcija antigena i limfocita
se dešava kada je:
- antigen prisutan u velikoj
koncentraciji u timusu
- afinitet receptora za antigen
visok
Centralna tolerancija se dešava i na
nivou autoreaktivnih CD4+ i CD8+ T
limfocita.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
isključivo za potrebe predavanja
www.belimantil.info
236.
PERIFERNA TOLERANCIJA TLIMFOCITA
Indukuje se kada zreli T limfociti prepoznaju autoantigene u
perifernom limfoidnom tkivu
Mehanizmi periferne tolerancije su:
1. Funkcionalna inaktivacija (anergija)
2. Smrt autoreaktivnih limfocita
3. Aktivna supresija autoreaktivnih limfocita
www.belimantil.info
237.
1. FUNKCIONALNA
INAKTIVACIJA
(ANERGIJA):
Dešava se
-kada zreli limfocit prepozna
antigen u odsustvu
kostimulacionih signala (drugi
signal aktivacije, npr.
interakcija B7:CD28), ili
- usled ekspresije inhibišućeg
receptora CTLA-4 na T
limfocitima ( interakcija
B7:CTLA-4 (CD152) generiše
inhibtorni signal)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanja
www.belimantil.info
238.
2. AKTIVACIJOM
INDUKOVANA SMRT
LIMFOCITA(DELECIJA)
- Ponavljana aktivacija zrelih T
limfocita sopstvenim antigenima
pokreće apoptozu i eliminaciju
(deleciju) autoreaktivnih ćelija.
- Ko-ekspresija receptora smrti
(Fas) i njegovog liganda (FasL) na
istoj ćeliji i interakcija Fas:FasL
pokreće aktivaciju kaspaza
(citosolni enzimi) koje indukuju
apoptozu.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman,
isključivo za potrebe predavanja
www.belimantil.info
239.
3. SUPRESIJA IMUNOG
ODGOVORA
-Prilikomreakcije sa
sopstvenim antigenom, neki
autoreaktivni T limfociti mogu
da se diferenciraju u
supresorske (regulatorne)
ćelije koje inhibiraju aktivaciju
drugih autoreaktivnih ćelija.
- Mehanizmi supresije ?
Neki T limfociiti mogu da
stvaraju TGF-β i IL-10 citokine
koji inhibiraju aktivaciju
limfocita, makrofaga.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo
za potrebe predavanja
www.belimantil.info
240.
TOLERANCIJA ILI IMUNIODGOVOR ?
1. Sopstveni antigeni su prisutni u timusu gde indukuju
centralnu toleranciju; mikrobni antigeni se transportuju i
koncentrišu u perifernim limfoidnim organima.
2. Sopstvene antigene prezentuju APC u odsustvu
nespecifičnog imuniteta i sekundarnih signala, što
indukuje anergiju; mikrobni antigeni stimulišu
mehanizme nespecifične imunosti, ekspresiju
kostimulatora, sekreciju citokina, što obezbeđuje
sekundarne signale, proliferaciju i diferencijaciju
efektorskih T limfocita.
3. Sopstveni antigeni su prisutni celog života i uzrokuju
ponavljane aktivacije T limfocita, odn. aktivacijom
indukovanu smrt limfocita; većina mikroorganizama se
brzo eliminiše posredstvom imunog odgovora i zato nije
u stanju da izazove smrt antigen-specifičnih limfocita.
www.belimantil.info
241.
TOLERANCIJA B LIMFOCITA
Sopstvenipolisaharidi, lipidi, nukleinske kiseline
(T-nezavisni antigeni) indukuju toleranciju B
limfocita; sopstveni proteini takođe mogu
indukovati toleranciju B limfocita
Mehanizmi i principi centralne i periferne
tolerancije B limfocita su slični kao kod T
limfocita
www.belimantil.info
242.
CENTRALNA
TOLERANCIJA B
LIMFOCITA
Kada nezreliB limfocit snažno
reaguje sa sopstvenim
antigenima u kosnoj srži B
limfociti
-umiru apoptozom (negativna
selekcija), ili
-Menjaju specifičnost
receptora reaktivisanjem
rekombinacije gena za L lance
Ig (reprogramiranje receptora)
Shema preuzeta iz knjige Basic Immunology, Abbas i
Lichtman, isključivo za potrebe predavanja
www.belimantil.info
243.
PERIFERNA
TOLERANCIJA B
LIMFOCITA
-Zreli Blimfocti koji
prepoznaju velike
koncentracije sopstvenih
antigena u perifernom
limfoidnom tkivu, postaju
anergični i ne mogu ponovo
da odgovore na taj antigen
-Nema odgovora B limfocita na
T-zavisne antigene ukoliko
izostane pomoć Th ćelija koje
su odsutne ili tolerantne
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanja
www.belimantil.info
244.
AUTOIMUNOST
Imuni odgovor protivsopstvennih (autologih)
antigena
1-2% populacije pati od autoimunih bolesti
Autoimunost nastaje usled izostanka autotolerancije:
- primarnih nedostataka imunog sistema
- abnormalnosti sopstvenih antigena i njihovog
prikazivanja limfocitima
Autoimunost se manifestuje produkcijom autoantitela i aktivacijom
autoreaktivnih T limfocita
Autoimune bolesti su najčešće heterogene i multifaktorske, sopstveni
antigeni koji pokreću autoimunu reakciju, kao i ciljne ćelije/molekuli su
često nepoznati, a bolest se često klinički manifestuje kasno posle
započinjanja autoimunske reakcije
www.belimantil.info
245.
Poremećaji centralne tolerancijemogu dovesti do abnormalne selekcije autoreaktivnih limfocita i njihovog
odlaska iz centralnih limfnih organa do ciljnih tkiva.
Poremećaji periferne tolerancije mogu dovesti do pojačane limfoidne reakcije na sopstveno tkivo-antigen.
Tkivne abnormalnosti (npr. inflamacija) mogu stimulisati ulazak limfocita u tkiva i prezentaciju autoantigena od
strane tkivnih APC.
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
246.
INFEKCIJE MOGU BITI
FAKTORRAZVOJA
AUTOIMUNE BOLESTI:
-Stimulacija lokalne nespecifične
imune reakcije, ekspresija
kostimulatora na APC, sekrecija
citokina: drugi signal aktivacije,
prekid anergije autoreaktivnih T
limfocita;
-Neki peptidni antigeni
mikroorganizama su slični
sopstvenim antigenima:
molekulska mimikrija (ukrštena
reaktivnost);
-Superantigeni porekla
mikroorganizama (oligoklonska
stimulacija T limfocita sa
identičnim VβTCR);
-Formiranje neoantigena kod
virusnih infekcija;
-Oštećenja tkiva usled infekcije i
oslobađanje sekvestriranih
antigena (npr., oko, testis).
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanja
www.belimantil.info
247.
Genetski faktori:
Mnoge autoimunebolesti povezane su sa ekspresijom
određenih gena, posebno MHC (HLA) lokusa: incidenca
bolesti označena je kao “relativni rizik”, iako HLA alel nije
uzrok bolesti.
Određeni MHC aleli mogu biti faktor rizika zato što:
. Nisu u stanju da prihvate sopstvene antigene pa je
poremećen proces negativne selekcije T limfocita;
. Peptidni autoantigeni prezentovani u sklopu tih MHC
alela nisu sposobni da stimulišu nastanak supresorskih
ćelija.
www.belimantil.info
248.
Neki primeri “relativnogrizika” oboljevanja od autoimunih bolesti i
ekspresije MHC alela:
Bolest HLA alel “Relativni rizik”
Ankilozirajući spondilitis B7 90-100 puta
Reumatoidni artritis DR4 4
Insulin-zavisni dijabetes DR3/DR4 25
melitus
Pemfigus vulgaris DR4 14
www.belimantil.info
249.
PODSETNIK
1. Imunološka tolerancijaje specifična nereaktivnost na neki antigen
indukovana kontaktom limfocita i tog antigena. Sve jedinke su
tolerantne (nereaktivne) na sopstvene antigene. Tolerancija se može
indukovati ako se neki antigen unosi u određenom obliku i na određeni
način u organizam; ovaj fenomen se koristi kao terapijska strategija za
autoimune bolesti, alergije i za sprečavanje odbacivanja transplantata.
2. Nastanak i održavanje tolerancije na sopstvene antigene
(autotolerancija) ostvaruje se mehanizmima centralne i periferne
tolerancije.
3. Centralna tolerancija se indukuje posle kontakta nezrelih limfocita i
sopstvenih antigena prisutnih u velikoj količini u centralnim limfnim
organima (timus, kosna srž); periferna tolerancija se indukuje posle
kontakta zrelih limfocita i sopstvenih antigena u perifernom limfnom
tkivu.
www.belimantil.info
250.
4. Centralna tolerancija(negativna selekcija) je rezultat
visokoafinitetnog vezivanja autoantigena za TCR, odnosno BCR
nezrelih T, odnosno B limfocita. Posledica je smrt limfocita tj.,
delecija autoreaktivnih klonova T i B ćelija, ili reprogramiranje BCR.
5. Periferna tolerancija T limfocita se indukuje putem sledećih
mehanizama:
- funkcionalna inaktivacija (anergija) usled nedostatka kostimulatora
ili ekspresije inhibitornih receptora;
- delecija (smrt apoptozom) usled ponavljanih kontakata sa
sopstvenim antigenom;
- supresija potencijalno štetnih T limfocita.
Periferna tolerancija B limfocita se najčešće indukuje usled izostanka
pomoći T limfocita.
6. Autoimune bolesti nastaju usled poremećaja autotolerancije. Faktori
koji su značajni za razvoj autoimunosti su:
- imunološke abnormalnosti
- genetski faktori
- abnormalnosti tkiva (infekcje)
www.belimantil.info
251.
IMUNI ODGOVOR PROTIVTUMORA
I TRANSPLANTATA
Tumorska imunologija
Transplantaciona imunologija
www.belimantil.info
252.
NEKE ZAJEDNIČKE KARAKTERISTIKE:
1.Imuni odgovor protiv ćelija koje nisu mikrobnog
porekla (ćelije tumora, transplantata)
2. Antigeni tumora, odn. Transplantata koji se
prepoznaju kao strani, mogu biti ispoljeni na bilo
kom tipu ćelija koje su maligno transformisane,
odn. Koje se transplantiraju.
3. Glavni mehanizam uništavanja tumorskih ćelija i
ćelija transplantata su CTL
www.belimantil.info
253.
1. Koje antigenetumora ili transplantata imuni sistem
prepoznaje kao strane ?
2. Kako imuni sistem prepoznaje i reaguje protiv tumora ili
transplantata ?
3. Na koji način je moguće modifikovati imuni odgovor u
cilju stimulacije odbacivanja tumora, odnosno inhibicije
odbacivanja transplantata ?
www.belimantil.info
254.
IMUNI ODGOVOR PROTIVTUMORA
Imunološki nadzor:
•Jedna od fizioloških uloga specifičnog imuniteta
kojom se sprečava razmnožavanje transformisanih
ćelija i uništavaju tumorske ćelije pre nego što se
formira tumor
•Razvoj tumora i njegov efekat na organizam ukazuju
da je imuni odgovor prema tumorima slab i da ga rast
tumora lako prevazilazi
www.belimantil.info
255.
TUMORSKI ANTIGENI
A. KojeT limfociti prepoznaju
kao strane:
- mutirani normalni proteini
ćelija (hemijski karcinogeni,
radijacija)
- Proizvodi mutiranih ili
translociranih onkogena ili
tumor-supresorskih gena
koji su verovatno uključeni
u proces maligne
transformacije
- Normalni proteini koji su
ispoljeni u velikoj količini ili
aberantno
- Proizvodi onkogenih virusa
B. Koje T limfociti ne
prepoznaju kao strane
- Glikolizivani sopstveni
proteini
- CEA, AFP
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
256.
Najvažnji mehanizam jeubijanje
tumorskih ćelija od strane CTL,
jer je većina tumorskih antigena
endogeno sintetisana (citosolni
proteini) i prikazana u kompleksu
sa MHC molekulima klase I.
Tumorske ćelije su sposobne da
prikažu kompleks tumorski
antigen/MHC I CD8+T limfocitima,
ali ne eksprimiraju kostimulatore;
za nastanak efektorskih CTL
neophodna je kostimulacija i/ili
pomoć specifičnih CD4+Th ćelija.
Profesionalne APC ingestiraju
tumorske ćelije i prezentuju
tumorske antigene u kompleksu
sa MHC I ili MHC II molekulima i
obezbeđuju kostimulaciju CD8+ i
CD4+ T limfocitima (ukrštena
prezentacija)
Kada uz pomoć antigena,
kostimulatora i Th nastanu
efektorski antitumorski CTL, oni
ubijaju tumorske ćelije bez dalje
potrebe za koostimulacijom ili Th.
Uloga antitumorskih antitela,
aktivisanih makrofaga i NK ćelija
IMUNOLOŠKI MEHANIZMI ODBACIVANJA TUMORA
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za
potrebe predavanja
www.belimantil.info
257.
TUMOR IZMIČE IMUNOM
ODGOVORU
A.Zato što je imuni odgovor
neefikasan:
- tumorski antigeni su najčešće
slabi imunogeni- hemijski se malo
razlikuju od normalnih sopstvenih
antigena.
B. Zato što tumor evoluira sa ciljem
da izbegne imuni odgovor:
- Tumor prestaje da ispoljava
tumorske antigene;
- Tumor prestaje da eksprimira
MHC I molekule
- Tumor proizvodi supresorske
molekule (npr., TGF-β)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
258.
IMUNOTERAPIJA TUMORA
Strategije imunostimulacije:
1.Pasivna imunizacija: unošenje tumor.specifičnih
antitela ili T limfocita
• Monklonska antitela protiv tumorskih
antigena
Efekat: toksični, citoliza, fagocitoza
Primer: anti-HER-2/neu u terapiji karcinoma dojke
mabtera anti-CD20 u terapiji B limfocitnog limfoma
• T limfociti izolovani iz krvi ili tumorskih
infiltrata bolesnika, stimulisani in vitro faktorima
rasta i vraćeni u organizam obolelog: adoptivni
transfer
www.belimantil.info
259.
2. Aktivna imunizacija(vakcinacija) tumorskim
antigenima:
• Tumorskim ćelijama ili tumorskim antigenima
izolovanim iz tumorskih ćelija
• Rekombinantnim proteinima sa adjuvansom
3. Stimulacija anti-tumorskog odgovora pomoću
specijalno pripremljenih DC ili tumorskih ćelija kao
vid vakcinacije
www.belimantil.info
260.
A. Izlacija DCiz krvi, stimulacija in vitro
faktorima rasta i tumorskim
ćelijama/antigenima i vraćanje
stimulisanih DC u organizam bolesnika.
B. Korišćenje plazmida koji sadrži cDNK
koji kodira tumorski antigen; unošenjem
ovakvog plazmida, cDNK se eksprimira u
ćelijama domaćina, uključujući i DC.
Ćelije domaćina proizvode tumorske
antigene i indukuju specifičan T ćelijski
odgovor.
C. U slučajevima kada je teška identifikacija
imunogenog tumorskog antigena, vrši se
stimlacija anti-tumorskog odgovora
pomoću:
- citokina (IL-2 i dr.);
- transfekcija tumorskih ćelija genima za
citokine (pr., IL-2) ili kostimulatore (pr.,
B7) i korišćenje transficiranih tumorskih
ćelija za vakcinaciju;
- eliminacija normalnih inhibitornih
signala imunog odgovora (pr., blokada
CTLA-4)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
261.
IMUNI ODGOVOR PROTIV
TRANSPLANTATA
Odbacivanjetransplantata (kalema) je imunološki
fenomen
Odbacivanje transplantata determinisano je
genima čiji su produkti ispoljeni na svim tkivima
Donor (davalac)/recipijent (primalac, domaćin)
Singeni transplantati (kalemi)
Alogeni antigeni
Ksenogeni antitela/T limfociti
www.belimantil.info
262.
TRANSPLANTACIONI ANTIGENI:
1. Imunološkareakcija u transplantaciji usmerena je
protiv MHC (HLA) molekula: transplantacioni antigeni
2. Svaka jedinka ispoljava 6 alela MHC I molekula (po
jedan HLA-A, B, C od svakog roditelja) i najmanje 6
alela MHC II molekula (HLA-DR, DP, DQ od svakog
roditelja) i njihove različite kombinacije,
3. MHC geni su visoko polimorfni: najmanje 120 alela
HLA-A i 250 alela HLA-B gena u populaciji.
4. Svaka individua ispoljava neke MHC molekule koji su
različiti od MHC molekula druge individue (izuzetak
jednojajčani blizanci).
5. Svi MHC molekuli mogu biti ciljevi odbacivanja, mada
su HLA-C i HLA-DP najmanje polimorfni i imaju manji
značaj.
6. I drugi polimorfni geniski proizvodi (koji nisu MHC)
mogu imati udeo u odbacivanju
www.belimantil.info
263.
A. T limfocitinormalno prepoznaju
kompleks strani peptid/sopstveni
MHC molekul.
B. Aloprepoznavanje MHC molekula
druge individue je jedan od
najsnažnijih imunih odgovora.
Direktno prepoznavanje alogenog
MHC molekula čija struktura
podseća na kompleks strani
peptid/sopstveni MHC molekul
(ukrštena reakcija). Donorov peptid
ne ušestvuje u aloprepoznavanju.
C. Aloprepoznavanje MHC molekula i
dela donorovog peptida
PREPOZNAVANJE
ALOGENIH MHC MOLEKULA
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanjawww.belimantil.info
264.
INDUKCIJA IMUNOG ODGOVORA
PROTIVTRANSPLANTATA
T limfociti mogu prepoznati
alogene MHC molekule
transplantata na različite načine,
zavisno od toga koje ćelije u
transplantatu prikazuju
aloantigene.
A. Ako transplantat sadrži profesionalne
APC koje ispoljavaju alogene MHC
molekule, to je direktna prezentacija,
odn., direktno prepoznavanje
aloantigena. Direktno prepoznavanje
stimuliše nastanak aloreaktivnih CTL.
B. Ako transplantat ne sadrži
profesionalne APC, verovatno je da
profesionalne APC primaoca ingestiraju
ćelije transplantata, aloantigeni se u
njima prerađuju i prezentuju u
kompleksu sa spstvenim MHC
molekulima; to je indirektna
prezentacija, odn., indirektno
prepoznavanje aloantigena. Indirektno
prepoznavanje stimuliše nastanak CTL
(ako je aloantigen u kompleksu sa
sopstvenim MHC I) ili CD4+Th ćelije
(ako je aloantigen u kompleksu sa
sopstvenim MHC II). CD4+ Th ćelije
sekretuju citokine koji utiču na
oštećenje transplantata po tipu
hroničnog odbacivanja (kasne
preosetljivosti)
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe
predavanja
www.belimantil.info
265.
IMUNOLOŠKI MEHANIZMI
ODBACIVANJA
TRANSPLANTATA
A. Hiperakutnoodbacivanje (minuti):
- cikulišuća, preformirana aloantitela
protiv antigena endotelnih ćelija
transplantata
- aktivacija komplementa i sistema
koagulacije
- tromboza krvnih sudova, ishemijska
nekroza
Prevencija: ukršteno sparivanje
Problemi: ksenotransplantacija
B. Akutno odbacivanje (dani.nedelje):
- CTL direktno ubijaju ćelije
transplantata, krvnih sudova
- antitela
C. Hronično odbacivanje (meseci-
godine)
- T limfociti, citokini
- hronična inflamacija, proliferacija
veziva (fibroza), arterioskleroza,
gubitak funkcije transplantata
Shema preuzeta iz knjige Basic Immunology, Abbas i Lichtman, isključivo za potrebe predavanjawww.belimantil.info
266.
PREVENCIJA I LEČENJEODBACIVANJA TRANSPLANTATA
Imunosupresija: inhibicija aktivacije i efektorske funkcije T
limfocita
Lek Mehanizam dejstva
Ciklosporin, FK506 Blokira produkciju IL-2 inhibicijom
aktivacijeNFAT transkripcionog faktora
Mikofenolat mofetil Blokira proliferaciju limfocita inhibicijom
sinteze guanin nukleotida
Rapamicin Blokira proliferaciju limfocita inhibicijom IL-
2 signala
Kortikosteroidi Redukcija inflamacije inhibicijom sekrecije
makrofagnih citokina
Anti CD3 monoklonskoantitelo Deplecija T ćelija
Anti IL-2R antitelo Inhibicija proliferacije T ćelija
CTLA-4-Ig Inhibicija aktivacije T ćelija bokiranjem
interakcije B7:CD28
Anti CD40L Inhibicija interakcije CD40:CD40L i
blokiranje stimulacije APC
www.belimantil.info
267.
TRANSPLANTACIJA KRVI IKOSTNE SRŽI:
Transplantacija ćelija krvi (transfuzja):
- ABO sistem krvnih grupa (antigeni A i B)
- Rh sistem krvnih grupa (C, D E, c, d, e antigeni)
Transplantacija kostne srži:
- da bi se korigovali defekti hematpoeze
(imunodeficijencije, autoimune bolesti);
- da bi se obnovila kostna srž posle zračenja ili
hemoterapije
www.belimantil.info
268.
TRANSPLANTACIJA KOSTNE SRŽI:
•Pretransplantacije kostne srži, nekad je potrebno uništiti kostnu
srž primaoca, kako bi se “oslobodio” postor za transplantat kostne
srži
•Imuni sistem reaguje snažno na alogene ćelije kostne srži, pa je
neophodno veoma precizno HLA sparivanje (tipizacija)
•Ako se uz alogene ćelije kostne srži transplantiraju i zreli T
limfociti davaoca, on mogu reagovati protiv primaočevih antigena i
dovesti do ozbiljnog kliničkog oštećenja poznatog kao reakcija
kalem-protiv-domaćina
www.belimantil.info






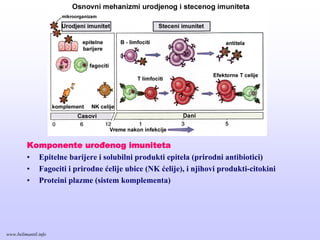
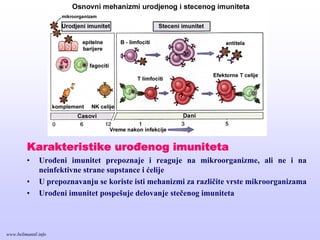
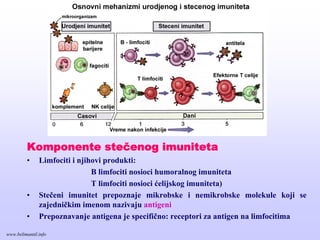
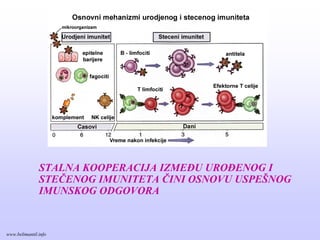


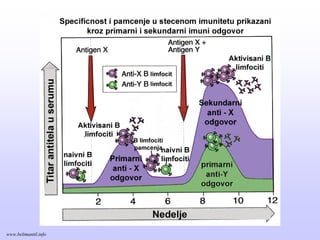
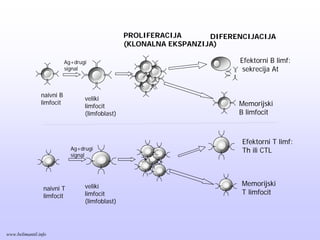
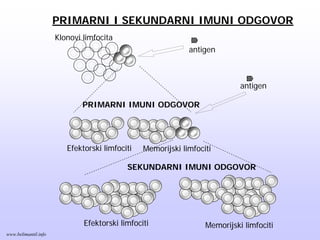

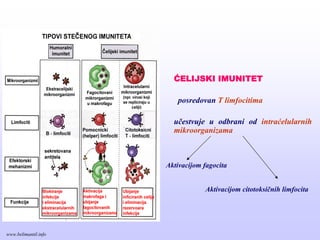

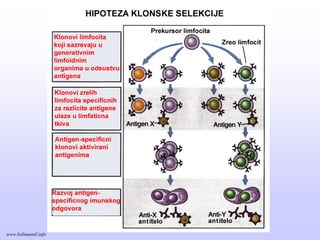
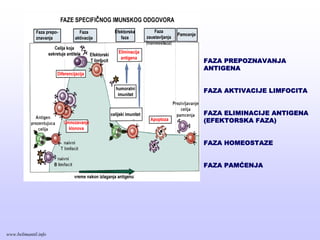


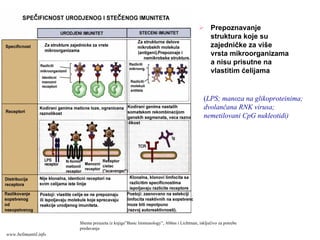
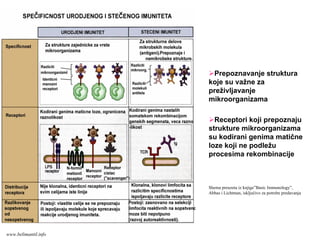

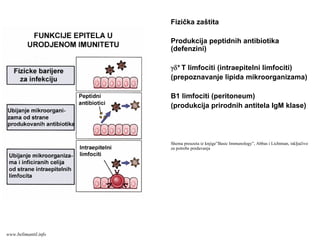
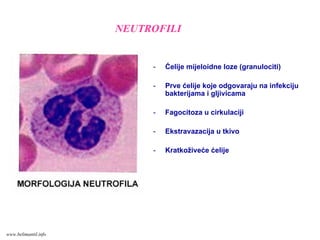
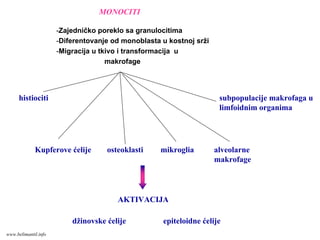
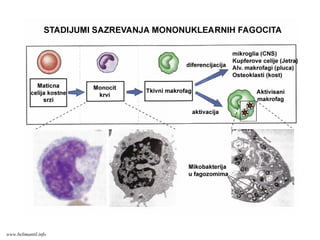
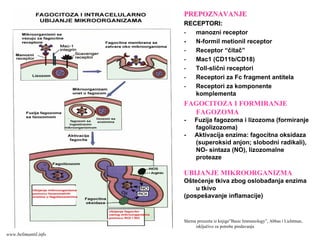
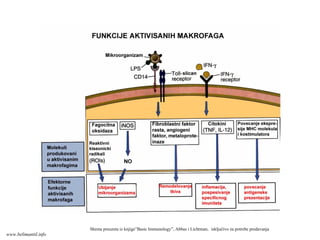
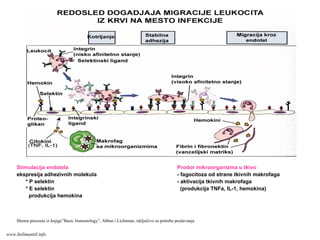
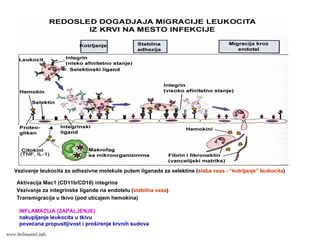
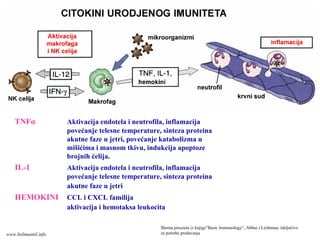
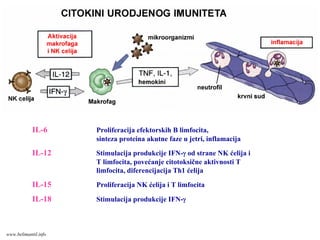
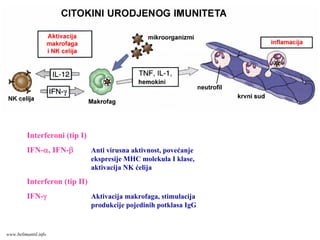
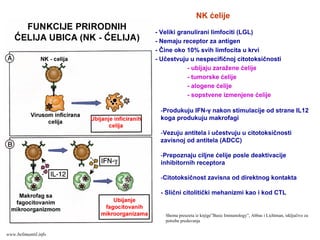
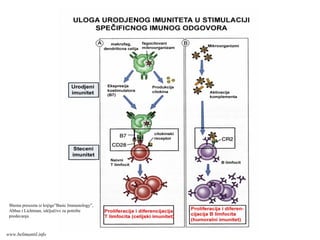








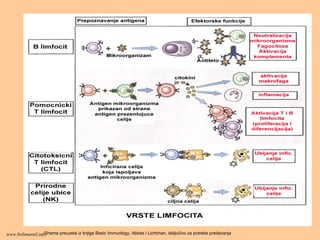

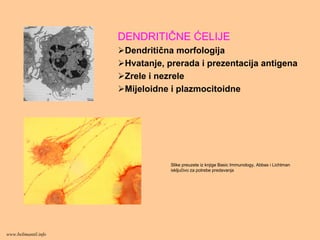

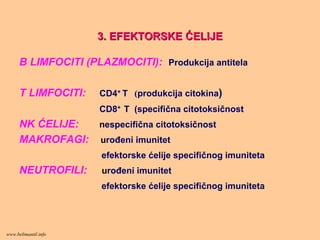

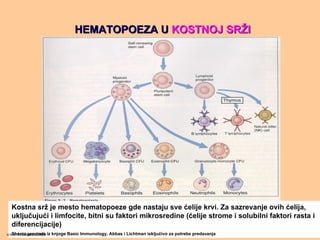

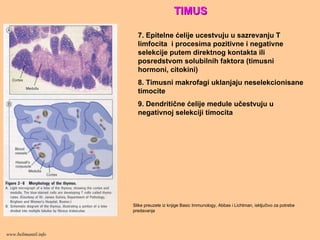
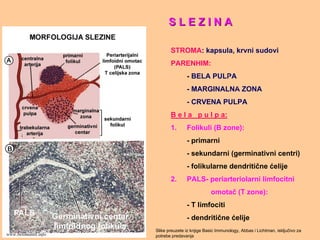
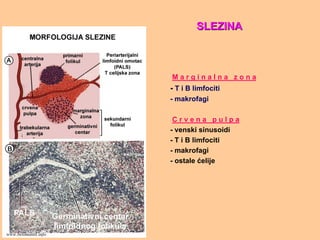
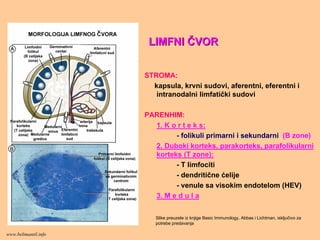
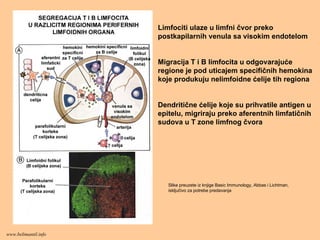
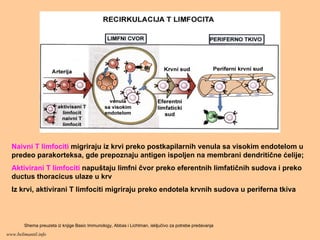






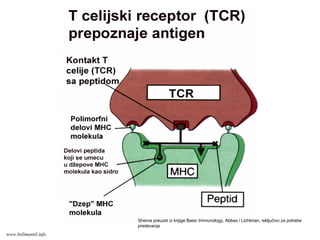



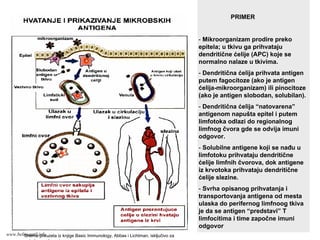
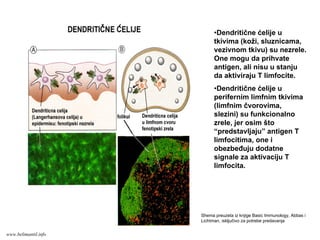
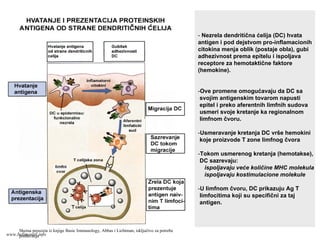


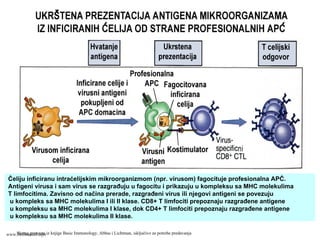



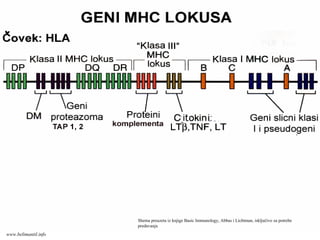


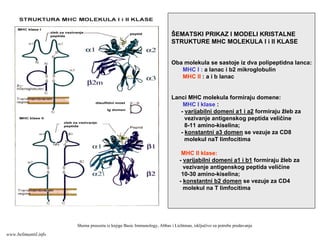

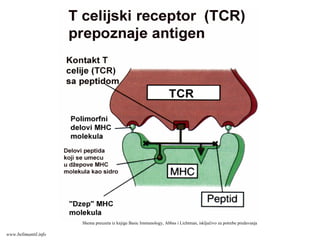



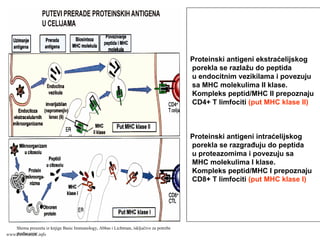
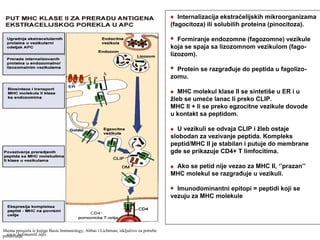
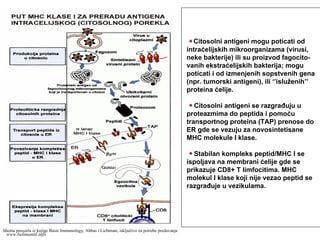
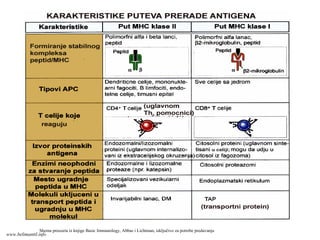


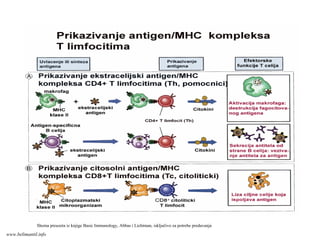





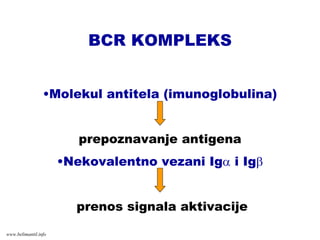
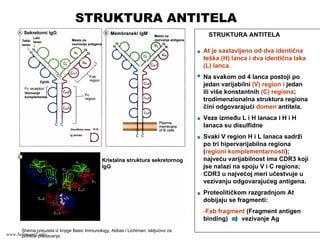
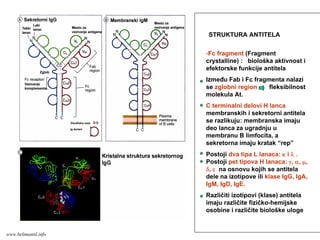
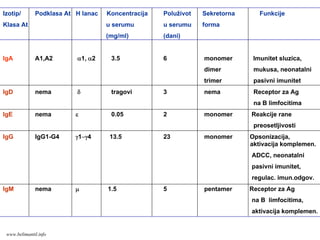

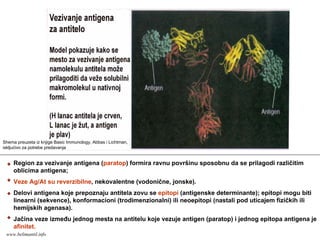
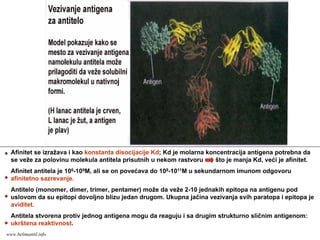

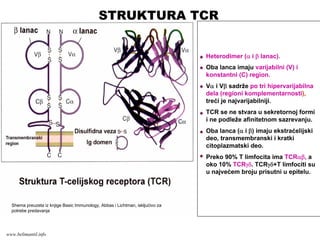

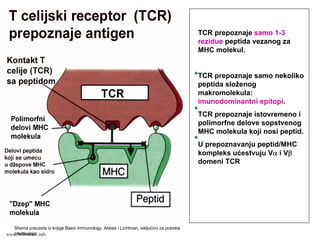
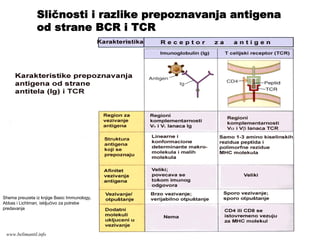


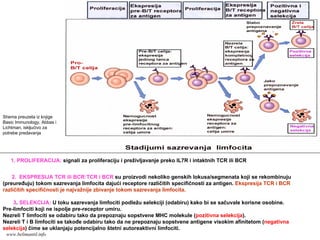


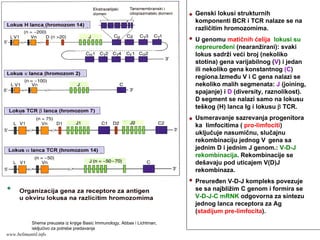
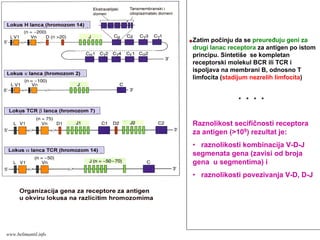
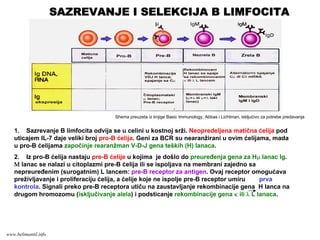
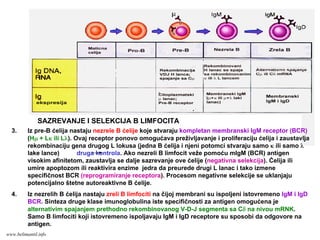
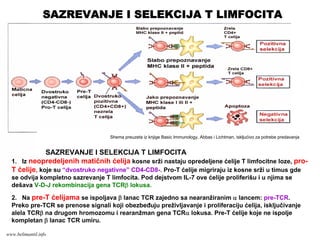
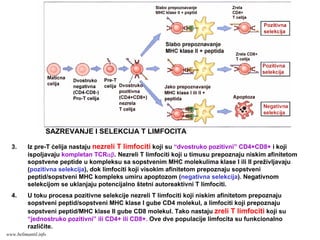






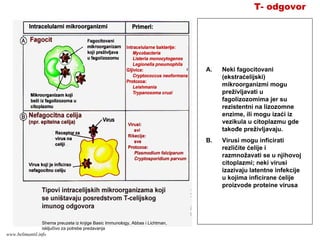
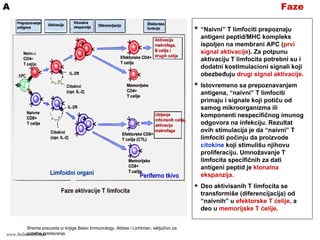
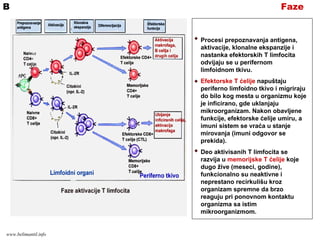
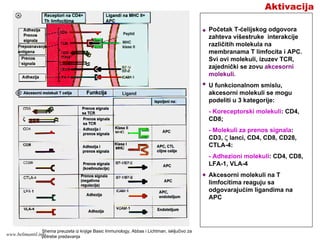
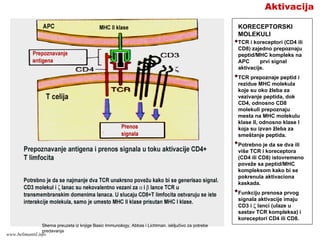

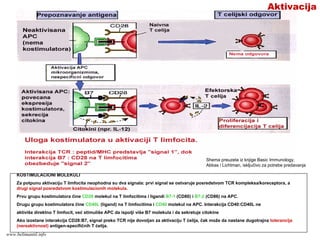
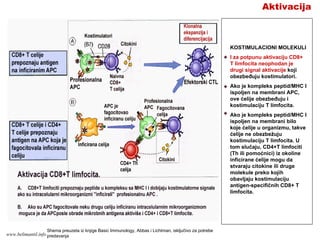

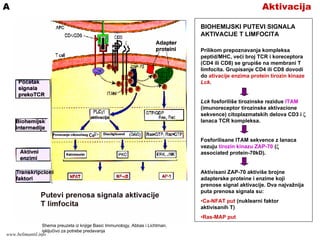
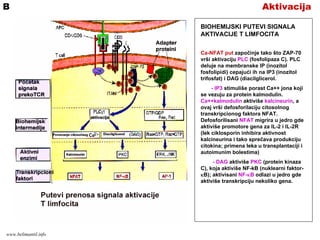
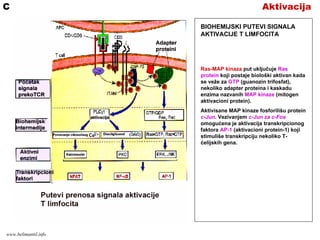

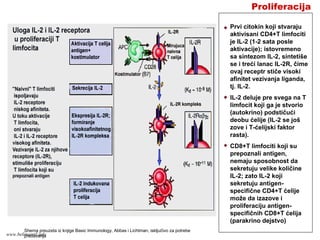


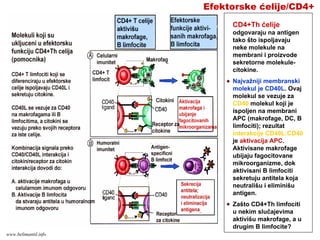
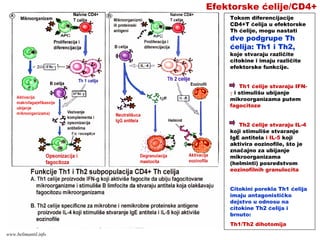
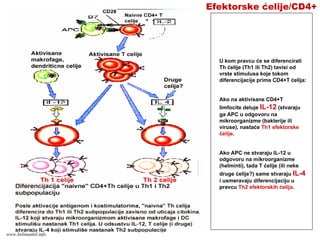


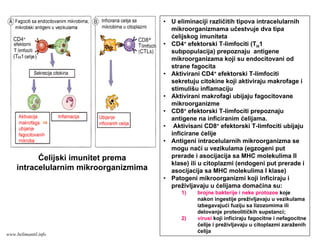
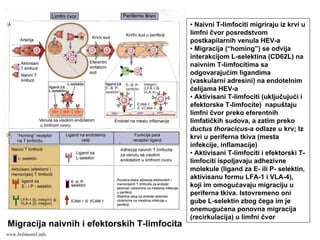
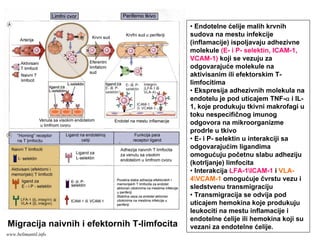
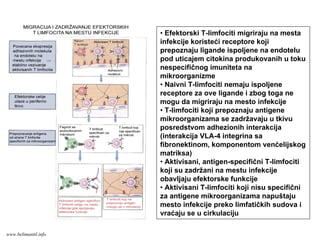

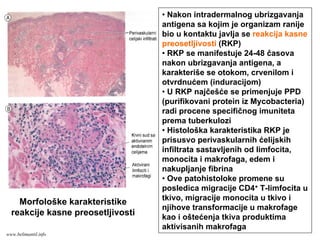

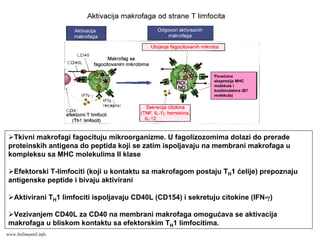
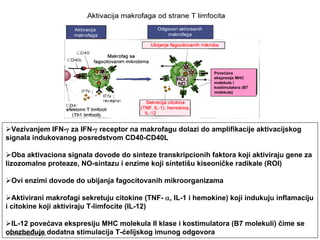


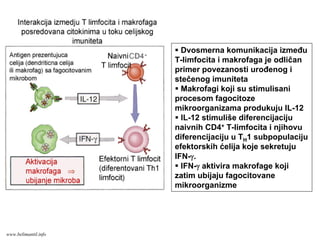
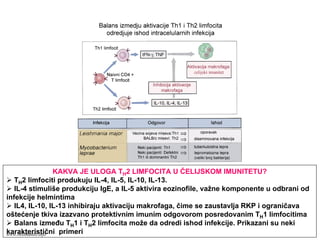

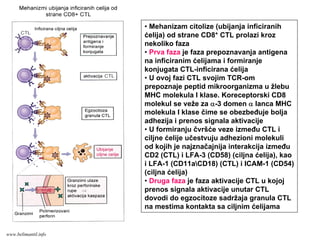
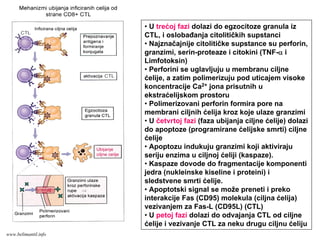
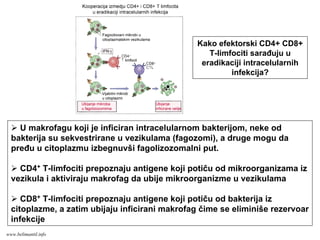





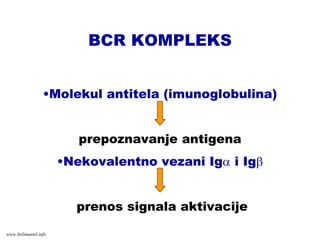
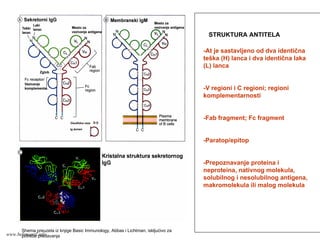

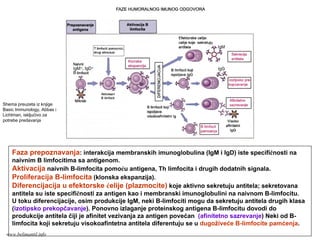

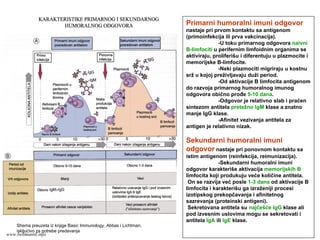
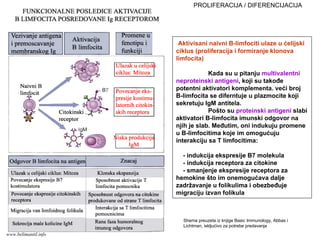

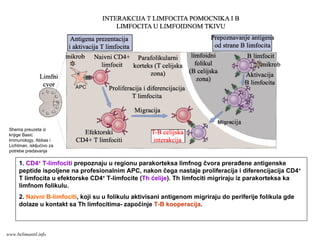
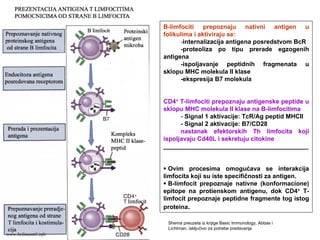
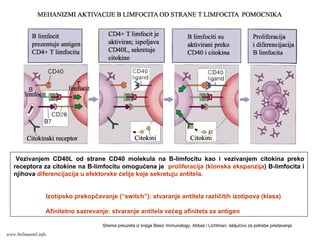
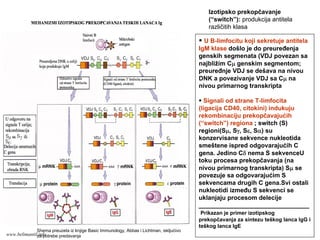
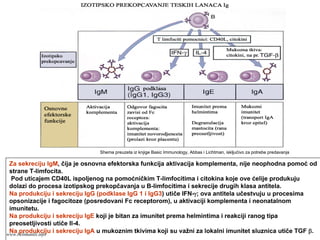
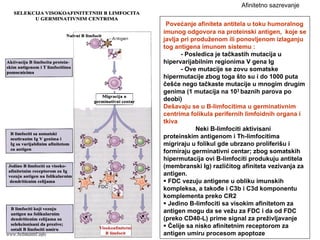
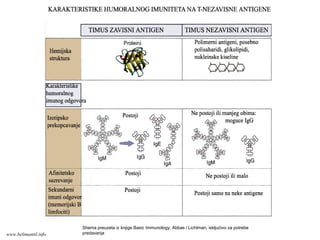
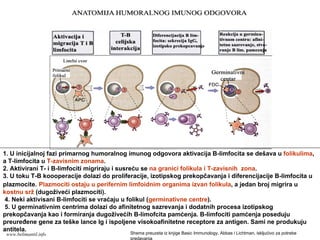



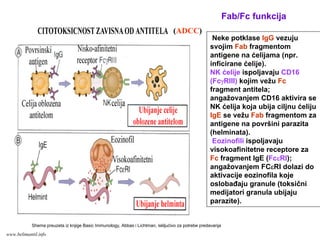
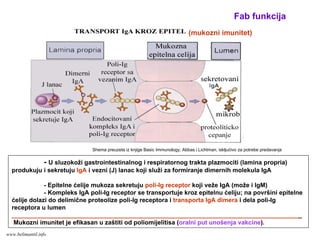


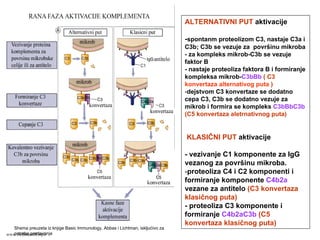
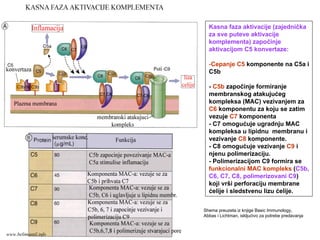
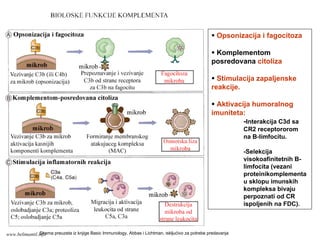
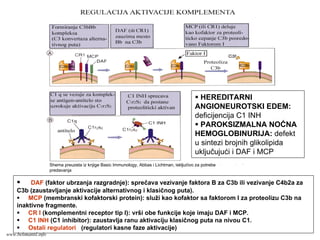
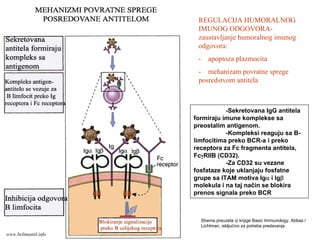



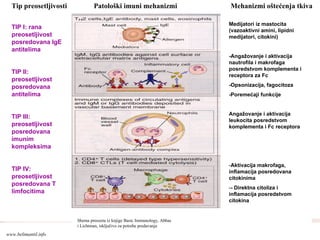


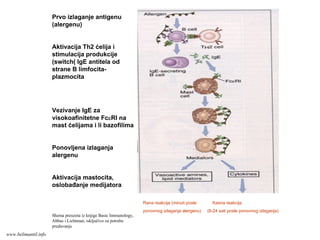
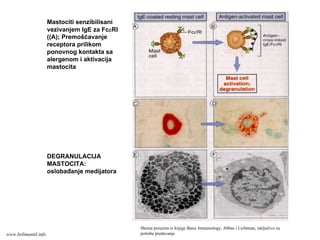

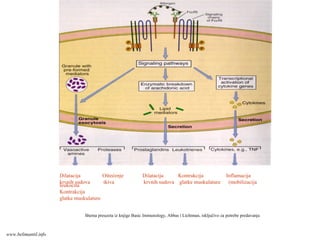


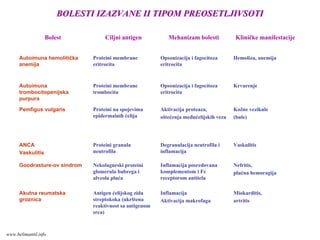
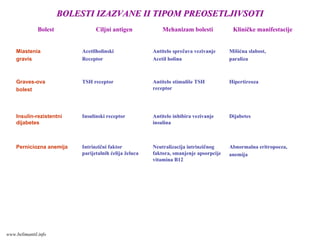


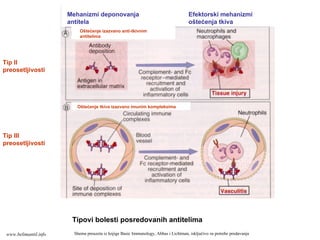
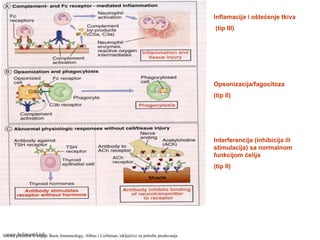

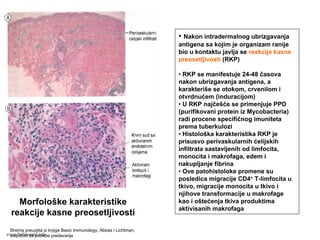

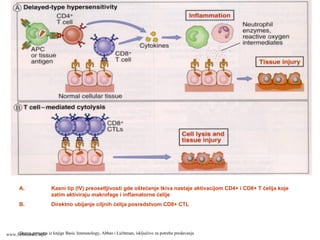



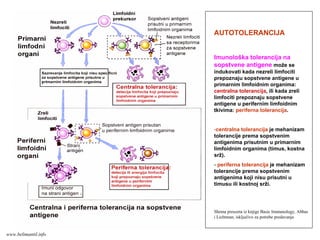
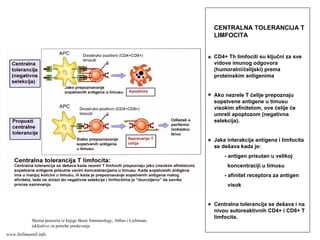

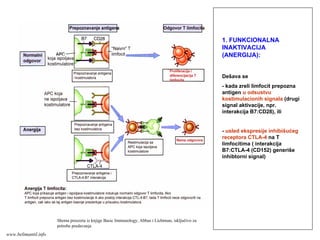
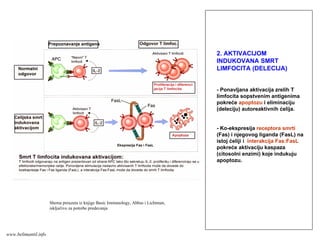
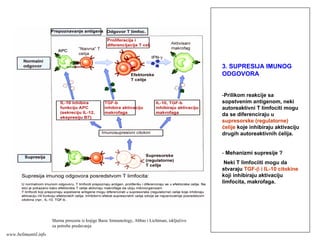


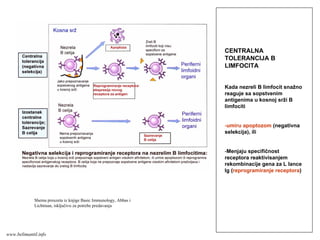
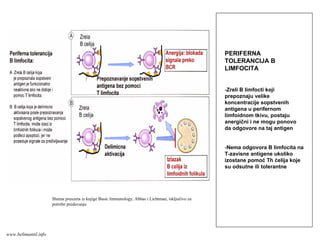

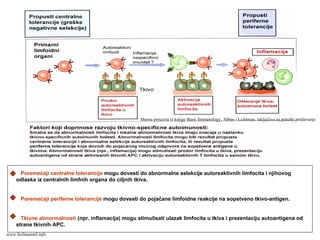

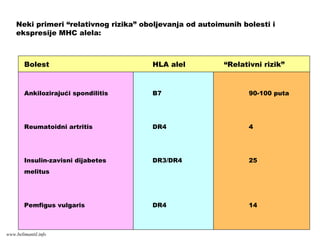






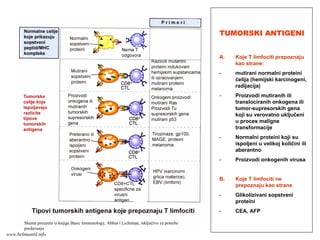
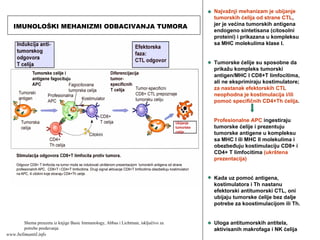
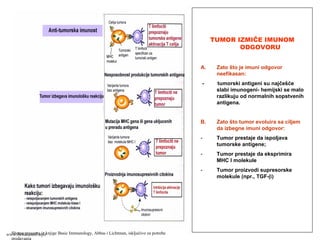


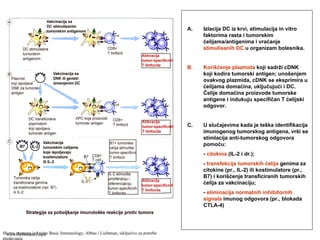
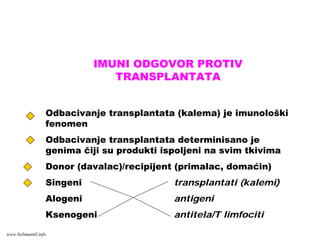

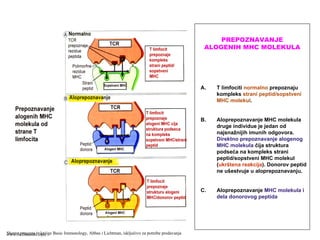
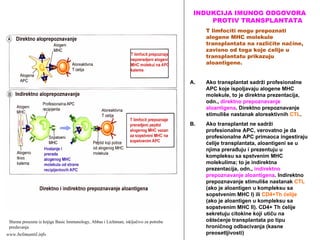
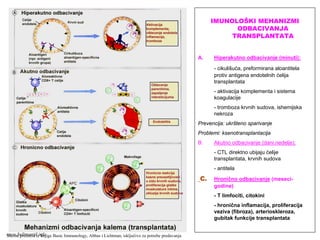
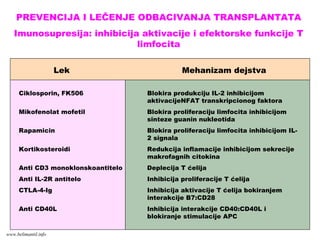















































































![Osnovi hirurgije Gerzic [pojedina poglavlja]](https://cdn.slidesharecdn.com/ss_thumbnails/osnovihirurgije-gerzicpojedinapoglavlja-141205055755-conversion-gate01-thumbnail.jpg?width=640&height=640&fit=bounds)








