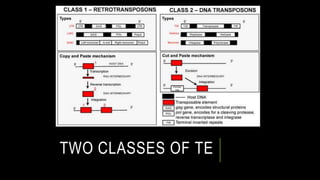
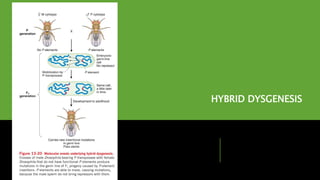
Transposons, or 'jumping genes', are DNA sequences that move within the genome, first discovered by Barbara McClintock in the 1940s. In Drosophila, P elements, a type of Class II transposable elements, can induce hybrid dysgenesis when crossed with M strain flies, leading to genetic defects and reduced fertility. The mechanism of transposition involves a protein called transposase, which is crucial for the activity of P elements, particularly in germline cells where specific splicing events occur.








































![63-79a-Chapter 2 PhD-Course. model organisms and their Genomes [Autosaved] (1...](https://cdn.slidesharecdn.com/ss_thumbnails/63-79a-chapter2phd-course-250324043930-2a63ecfb-thumbnail.jpg?width=640&height=640&fit=bounds)





































