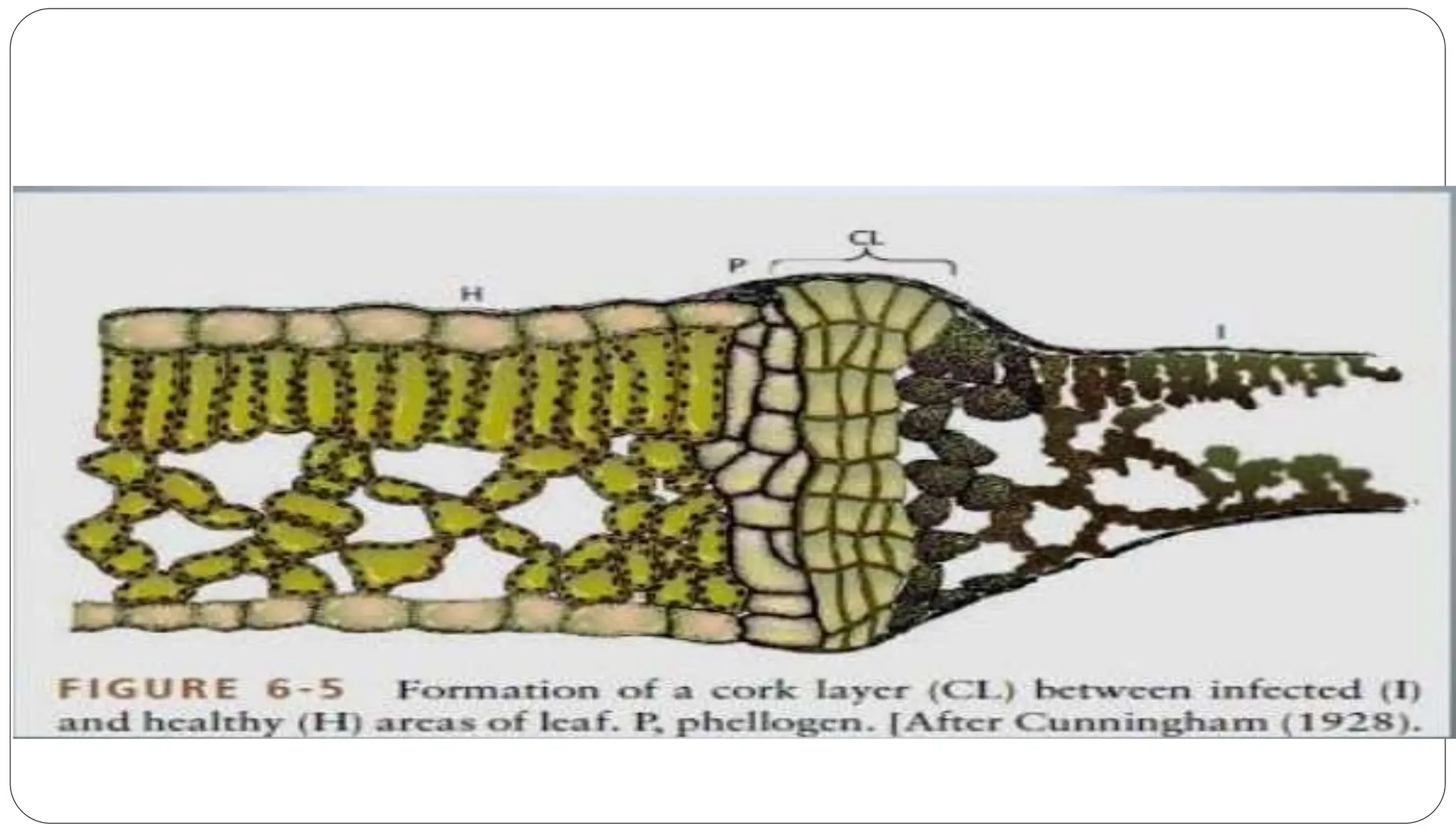
Plants have two types of defense mechanisms against pathogens: structural barriers and biochemical reactions. Structural barriers include pre-existing features like thick cell walls and wax coatings, as well as post-infection responses like cork layers, tyloses, and necrotic tissue that isolate the pathogen. Biochemical defenses include pre-existing inhibitors exuded by the plant and induced responses like the hypersensitive response where infected cells undergo rapid cell death to prevent pathogen spread. Together these mechanisms inhibit pathogens from entering plants, limit their growth and movement within plants, and produce toxic compounds to kill pathogens.