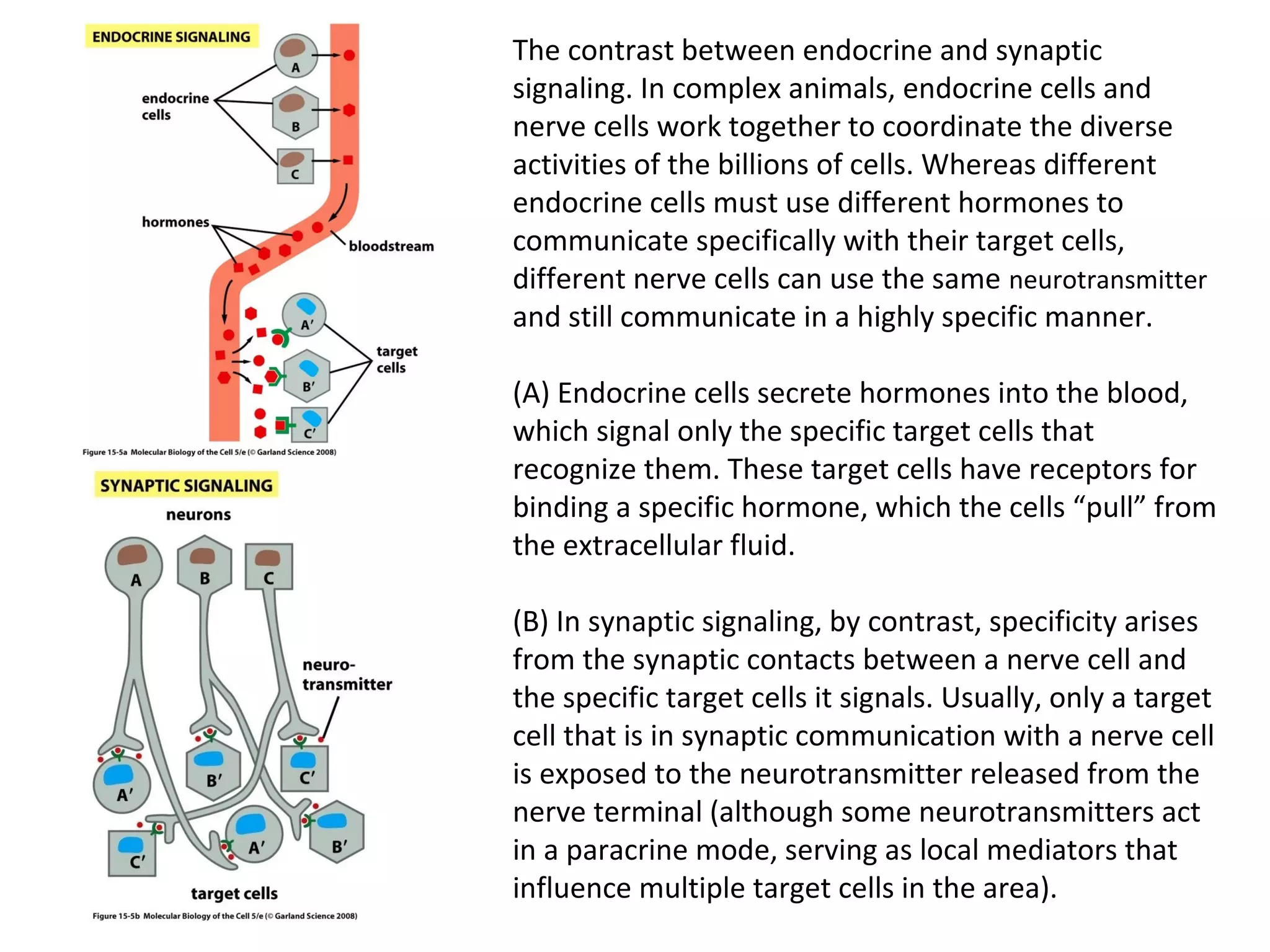
The document discusses cell signaling mechanisms essential for multicellular organisms, highlighting how cells communicate via extracellular signal molecules. It explains various signaling types, such as paracrine, synaptic, and endocrine signaling, and emphasizes the complexity of receptor interactions and the necessity for multiple signals for cellular functions. Additionally, it describes the roles of nuclear hormone receptors and the integration of signals through intracellular pathways to regulate cell behavior.