Downloaded 130 times




















![Objective :To understand sex biasness in the absence of ecological disturbance
in dioecious species ‘Salix viminalis’ .
Experimental Material - Salix viminalis [Basket willow]
These are multi stemmed shrub growing to between 3 and 6 m.
Male and female catkins are borne on separate plants.
Commonly used in basketry. Other uses: effluent treatment in wastewater
gardens, and for water purification.
Case Study-1](https://image.slidesharecdn.com/sexdeter-180816141127/75/SEX-DETERMINATION-MECHANISMS-IN-PLANTS-22-2048.jpg)

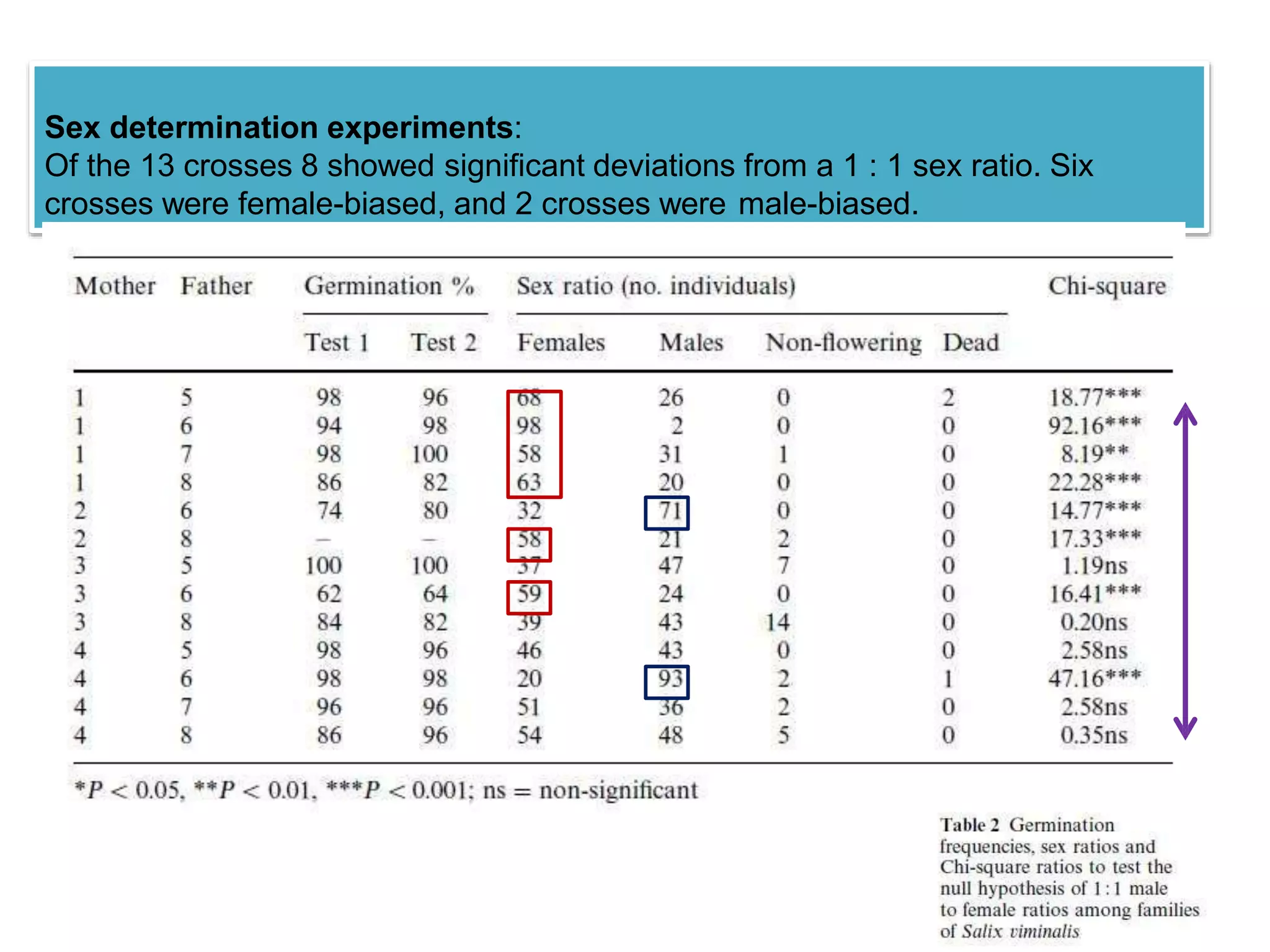
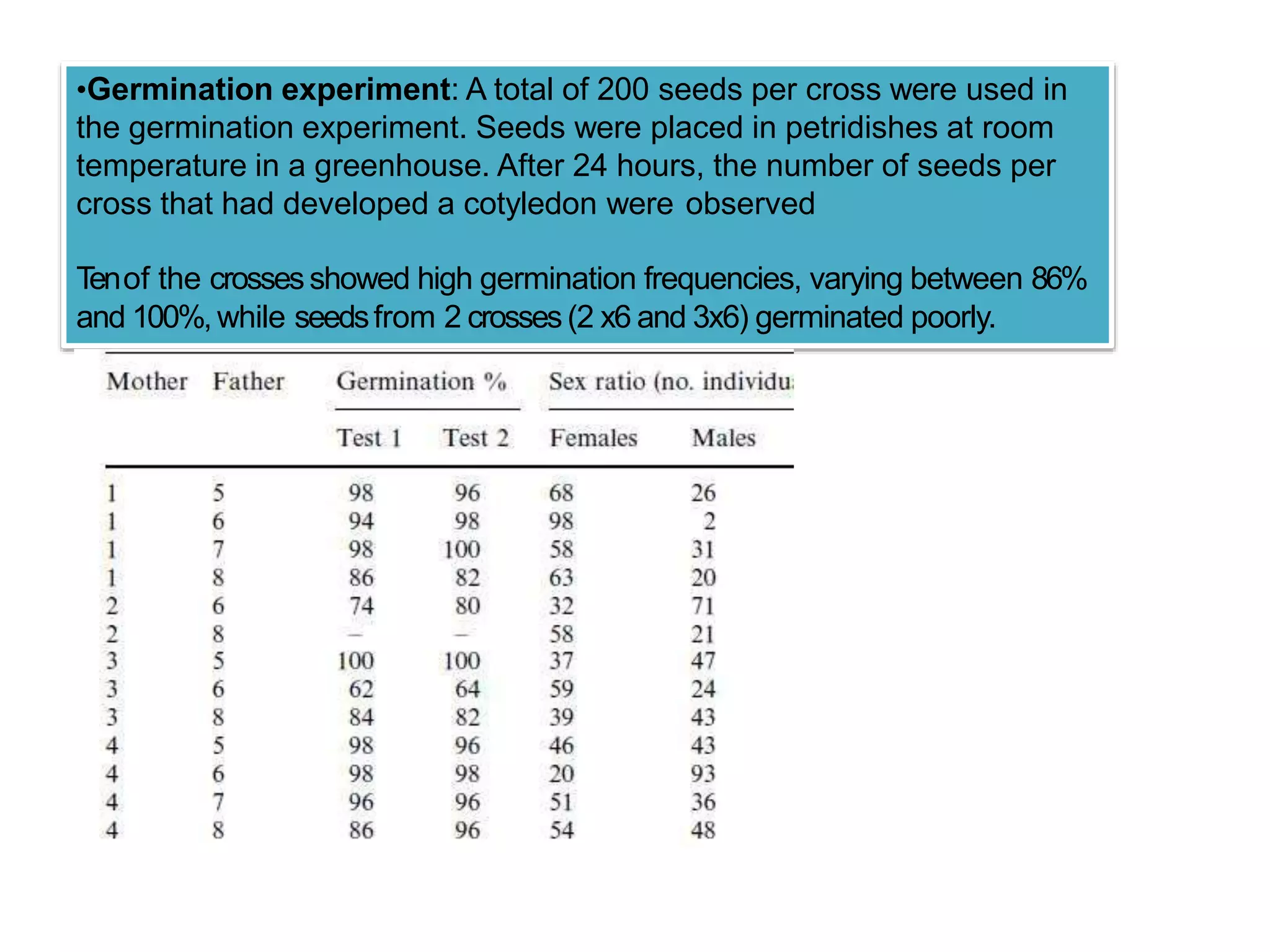














This study investigated sex determination in the dioecious plant species Salix viminalis (basket willow). Crosses between 4 female and 4 male parents resulted in 13 offspring crosses, of which 6 were female-biased and 2 were male-biased in their sex ratios. A germination experiment found most crosses had high germination rates, indicating fitness differences did not cause the biased sex ratios. As no hermaphrodites or sex changing plants were observed, sexual lability also did not explain the biases. Meiotic drive or gametic selection could potentially cause the variation, but were deemed unlikely as biases existed among both same-father and same-mother crosses. The skewed ratios were also not due to cytoplas
Overview of biological sex-determination systems, genetic implications, and historical references.
Discusses evolutionary advantages of sex, including genetic variation and adaptation.
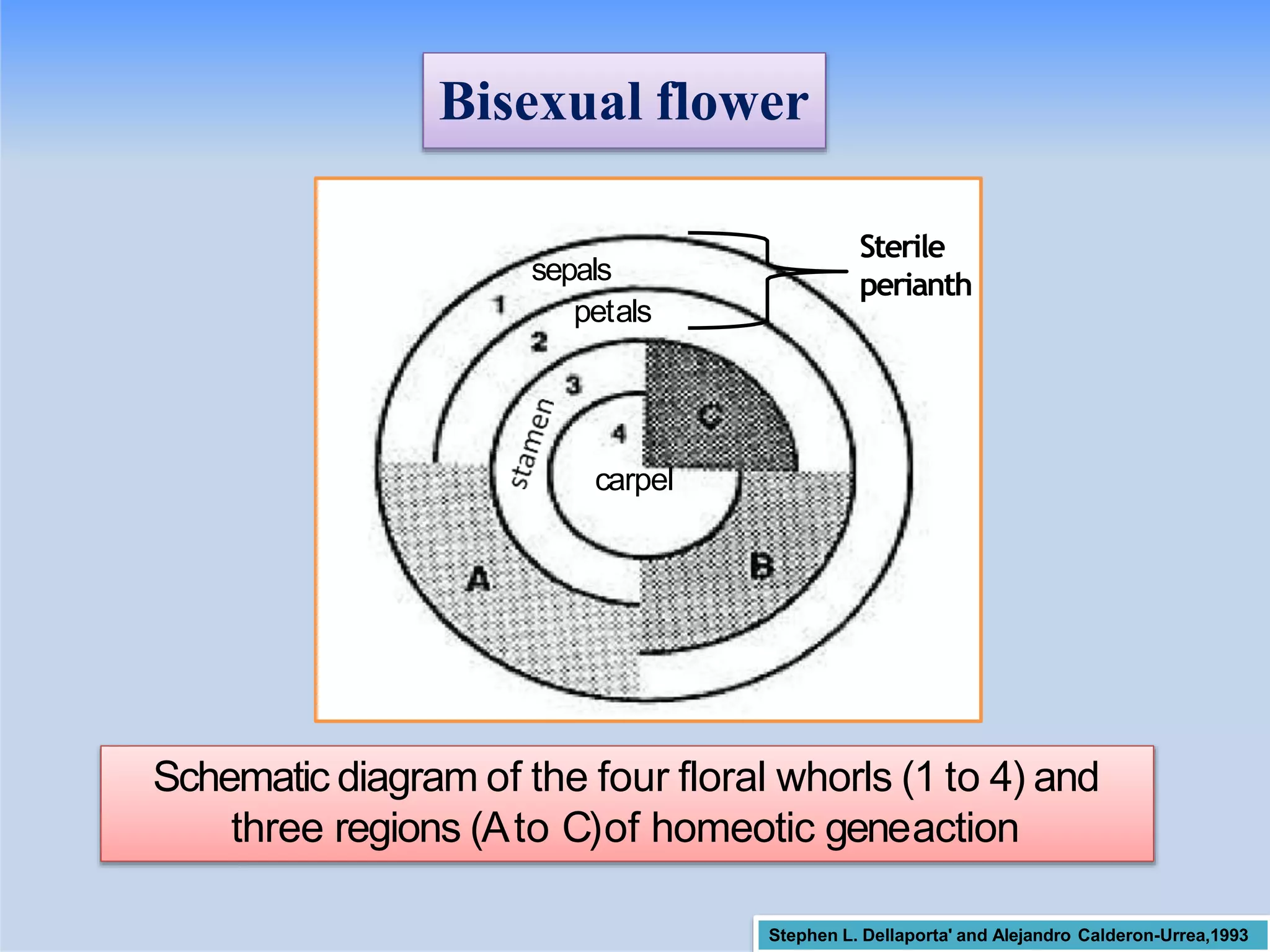
Illustrates different modes of sexuality in flowering plants that contribute to reproduction.
Explains various mechanisms like environmental, chromosomal, and genic factors in sex determination.
Describes how environmental conditions influence the sex determination in various plants.
Chronicles discoveries related to X and Y chromosomes and their roles in sex determination.
Distinguishes heteromorphic from homomorphic sex chromosomes and their genetic implications.
Examines sex determination governed by single or multiple gene loci in certain plants.
Outlines the impact of sex determination on flower development in maize and related species.Details an experiment on sex ratios in Salix viminalis with methods and findings regarding skewed ratios.
Explains methodology for determining sex in F2 papaya plants utilizing RAPD markers.Describes the results of sex determination in Simarouba glauca using RAPD markers.
Summarizes complexity of sex determination in plants and the importance of molecular markers for breeding.























































































