Downloaded 182 times



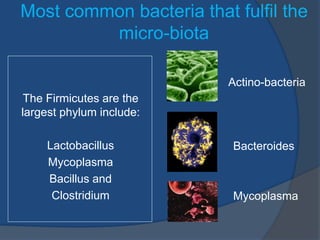
























- The human gut microbiota plays an important role in metabolism and energy homeostasis. It contains over 1000 bacterial species totaling 1014 bacteria. - Studies show the gut microbiota influences the development of type 2 diabetes by regulating peripheral metabolism and energy production. Variations in the gut microbiota can positively or negatively impact diabetes risk. - The gut microbiota matures during the first years of life and is influenced by factors like birth method, infant feeding, and use of antibiotics. Manipulating the gut microbiota through probiotics or prebiotics may help regulate glucose levels and reduce diabetes risk.









































































