This document discusses genetic processes that can change gene frequencies in populations. There are two main types:
1. Systematic processes like migration, mutation, and selection that tend to change gene frequencies predictably in amount and direction. They act in both large and small populations.
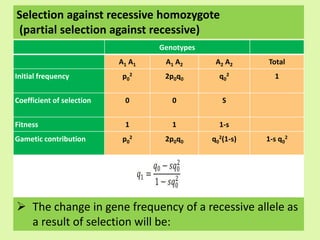
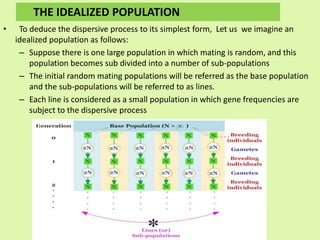
2. Dispersive processes that arise in small populations from random sampling effects. They are predictable in amount but not direction and only act in small populations. Random genetic drift is a main dispersive process.