CALL ON ➥8923113531 🔝Call Girls Kesar Bagh Lucknow best Night Fun service 🪡
Basic plant cytology
1. Plant Physiology
1
Basic Plant Cytology
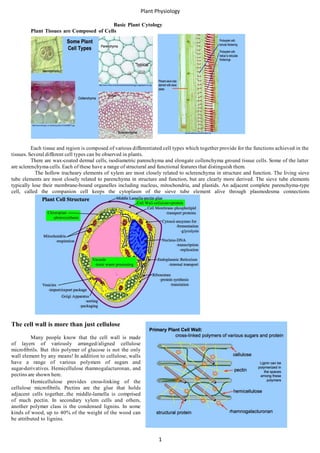
Plant Tissues are Composed of Cells
Each tissue and region is composed of various differentiated cell types which together provide for the functions achieved in the
tissues. Several different cell types can be observed in plants.
There are wax-coated dermal cells, isodiametric parenchyma and elongate collenchyma ground tissue cells. Some of the latter
are sclerenchyma cells. Each of these have a range of structural and functional features that distinguish them.
The hollow tracheary elements of xylem are most closely related to sclerenchyma in structure and function. The living sieve
tube elements are most closely related to parenchyma in structure and function, but are clearly more derived. The sieve tube elements
typically lose their membrane-bound organelles including nucleus, mitochondria, and plastids. An adjacent complete parenchyma-type
cell, called the companion cell keeps the cytoplasm of the sieve tube element alive through plasmodesma connections
The cell wall is more than just cellulose
Many people know that the cell wall is made
of layers of variously arranged/aligned cellulose
microfibrils. But this polymer of glucose is not the only
wall element by any means! In addition to cellulose, walls
have a range of various polymers of sugars and
sugar-derivatives. Hemicellulose rhamnogalacturonan, and
pectins are shown here.
Hemicellulose provides cross-linking of the
cellulose microfibrils. Pectins are the glue that holds
adjacent cells together...the middle-lamella is comprised
of much pectin. In secondary xylem cells and others,
another polymer class is the condensed lignins. In some
kinds of wood, up to 40% of the weight of the wood can
be attributed to lignins.
2. Plant Physiology
2
The cell wall possesses catalytic activity
But the cell wall is even more than just polysaccharide relatives. A critical component of cell walls is protein that
provides catalytic activity for the cell wall region. Enzymes that polymerize wall monomers, enzymes that cross-link polymers, enzymes
that cleave polymers, these and others permit the cell wall region to be a dynamically-sculpted element for a living cell.
The cell wall provides for turgor pressure
Far from being a barrier, the wall is partially for the structural support of a multicellular, multidimensional plant body, but has a
critical function in providing a means to survive in a dilute solution of solutes. Soil water is a hypotonic medium and the wall provides
a means to avoid cell expansion that would otherwise exceed the bursting strength of the cell membrane. It permits the development
of turgor pressure which can be a structural and functional factor in support and movement.
The cell membrane is more than phospholipids
The cell membrane, generally just inside of the cell wall and tightly apressed against it because of turgor pressure, is the
exchange regulator for the cell and its environment. Indeed the oligosaccharide subunits and wall-sculpting enzymes are passed from the
interior of the cell through this membrane: by exocytosis. Water, minerals, sometimes organic particles, pass from the environment
through this membrane to the cell's interior: endocytosis. While "goodies" are allowed to pass through the membrane, other
substances are kept outside. The cell membrane is shown below.
The phosopholipids provide barrier functions
The main barrier function is provided by the phospholipid bilayer. This bilayer is composed of amphipathic phospholipid
molecules. Two C14-C24 fatty acids, one saturated (no double bonds) and
the other at least mono-unsaturated (one double bond), comprise the
hydrophobic "tails" that make passage of charged and polar molecules
into and out of the cell all but impossible. These are linked by a glycerol
(3C-sugar) to a very polar and hydrophilic phosphate-small-organic-group
"head." Thus polar solutes might pass into the "heads" part of the
membrane, but cannot pass through the non-polar "tails." Similarly
non-polar molecules would pass easily through the "tails" part of the
membrane, but cannot approach that because of the polar "heads" layer.
Phospholipids spontaneously form bilayers with the polar "heads"
facing the aqueous extra-cellular fluids and the aqueous cytosol. Between
these layers of "heads" the hydrophobic "tails" inter-mingle. The double-layer
of "tails" thus provides a the barrier functions for many
biologically-interesting ions and molecules.An individual phospholipid in the
bilayer is free to move about in the plane of the bilayer. Contributing to this
fluidity of movement, or stabilizing this fluidity depending on temperature are various steroids that are often a part of the membrane. In
plants the steroid components include wide range of distinct molecules.
The cell membrane proteins provide catalytic functions
But a cell membrane that is a great barrier is not good for the cell per se. In fact a barrier is only as good as its doors and
windows. The mark of a good barrier is that it allows for exchange of positive elements while providing only exit and never entrance
of negative elements. This lesson, hopefully recalling the Iron Curtain, is pointing to the proteins of a cell membrane.
The phospholipid bilayer is traversed by integral proteins. These proteins have two hydrophilic zones separated by a
hydrophobic zone of suitable dimensions to become trans-membrane transport proteins. These may permit facilitated diffusion of
suitably small charged molecules or elements, or perhaps even active transport of essential components. Active transport of course
involves the conversion of ATP into ADP and Pi or AMP, with the released energy from the cleavage of the phosphate bond driving
the movement of the component through the membrane.
3. Plant Physiology
3
The membrane is also a site of catalytic activity.
In addition to integral proteins, there may also be
peripheral proteins attached to one side or the other of the
bilayer. The attachments may involve simple hydrophobic
and hydrophilic domains of the protein, but might also
involve linkage of the protein to fatty acids or other
hydrophobic attachments that anchor them within the
bilayer and leaving the protein facing the aqueous
environment. These proteins likely serve various catalytic
or electronic functions. Depending on which face of the
bilayer we are examining, we might find ATP synthases
or electron transfer proteins here. We might also find
enzymes, such as succinate dehydrogenase, that catalyze
steps associated with pathways in one compartment or
another adjacent to that surface of the bilayer. The many
proteins in a membrane may constitute 40% of that
membrane! To simply call a membrane a "phospholipid
barrier" would be a gross over-simplification.
The cytosol is more than just water
Inside the cell wall and membrane we have the fluid compartment of the cell. Indeed this compartment is mostly water...and
water has some very important roles to play in the life of a cell...but this fluid is much more than just water!
Saying "just water" is really an injustice to this critical molecule for life! We will get into that later in the course in some detail. Let's
remember it as the medium for the creation of life, as a UV screen until the ozone layer helped out, and as more than 90% of the weight
of metabolically active cells.
This region was called cytoplasm (literally cell fluid) back in the days when light microscopy was a cutting-edge
technology. When electron microscopy was developed, we learned that this cytoplasm was not just fluid...it contained
previously-invisible structures that are not simply fluid. So we use cytoplasm generally to mean the fluid and all of these
contained organelles together. A newer word, cytosol (literally cell solvent), is used to mean only the fluid in which the
organelles are suspended.
The cytosol hosts tremendous catalytic activity
The cytosol contains dissolved minerals, gases, and organic molecules. These provide for the many catalytic functions of the
cytosol. Dissolved proteins, often called soluble enzymes, are responsible for a range of biochemical sytheses and degradations that
characterize a living cell. The cytosol is host to a range of entire pathways of regulated biochemistry! Examples include
glycolysis, fermentation, and (from a certain perspective) translation!
In addition to these functions, the cytosol serves as a hydraulic fluid for organismal support, as a medium for diffusion, and as a
compartment for osmosis. It provides thermal buffering capacity and as a transparent medium for light penetration. The differential
solubility of oxygen gas and carbon-dioxide gas provides for a medium in which photosynthesis can occur. Dissolved pigments may
provide protection in times of excessive light irradiation.
The nucleus is more than just a DNA container
The nucleus is an organelle of the cell that is not considered part of the term cytoplasm. It was obvious in the early days of
microscopy. Various staining procedures revealed that it contained nucleic acids (DNA and RNA) well before the hereditary roles of
these molecules was known. But early tests also indicated the presence of proteins.
The nuclear envelope is composed of two bilayer membranes
The outer membrane and inner membrane of the nuclear envelope are separated by a perinuclear space. The bilayer facing
the cytosol is perhaps very much like that of the endoplasmic reticulum and often hosts polyribosomes. The bilayer facing the
nucleoplasm is coated with the nuclear lamina...a layer of intermediate filaments. During mitosis the envelope is dispersed as small
vesicles that coalesce after mitoses.
4. Plant Physiology
4
The nuclear envelope has "pores" for the passage of large materials
The nuclear pore complex is a formation of structural and functional proteins that permit the movement of molecules,
macromolecules, and even the subunits of ribosomes through the envelope. The pore complex is depicted below. A key element of this
complex is the transporter protein, which can be regulated to permit or prohibit movement through the envelope.
DNA in the nucleus is found in chromosomes
The hereditary molecule, DNA, is amazingly long. Each
molecule, called a chromosome, is composed of many shorter lengths
of DNA, called genes, that are linked end to end. The sequence of
nucleotides in each gene constitute the instructions for the synthesis of
a single protein. For various portions at one time or another, and for the
whole chromosome at the start of mitosis, the chromosome is condensed
in a process shown below.
Histone proteins are a primitive feature of most eukaryotic
organisms. The amino acid sequence of these proteins is exceedingly
conservative among plants, animals, and fungi. The DNA molecule is
wrapped around histones in the form of nucleosomes. These are
cross-linked by other histones, and coiled and looped in repeatable
ways to ultimately produce the familiar X-shaped chromosomes we
observe during mitosis. Because the set of n chromosomes, as a group,
represent one entire genome, and two such sets are present in most plant
nuclei, it is no surprise that most of the DNA in any particular cell is
not being expressed (used) at any point in time. When one considers
that each cell contains two complete sets of instructions for a whole organism, it is clear that most of the DNA must not be "active."
The nucleus often demonstrates a nucleolus
Early light microscope preparations were stained and even in interphase, when the chromosomes are NOT condensed, one or
more intensely-staining zones were observed in the nucleus. These were called nucleoli. RNA-specific stains and probes reveal that the
nucleolus is a region with much RNA compared to the rest of the nucleus. The nucleoli are considered to represent the location of
genes coding for ribosomal and transfer RNAs. In metabolically active interphase cells, many ribosomes and transfer RNAs are
needed to maintain the protein pools needed for active metabolism. These genes, then, are being transcribed at a very high rate and the
RNA products are locally abundant, explaining the intense local staining.
Protein stains also highlight the nucleolus. Proteins are translated in the cytosol and those destined for the nucleus pass
through the pore complex into the nucleoplasm.
Some of these are enzymes are involved in
replication and transcription, including DNA and
RNA polymerase. Other proteins arriving are the
components that must join ribosomal RNA to
form ribosome subunits. The two ribosome
subunits, the large and small subunits, are
assembled in the nucleolus region explaining the
protein stain results. The ribosome subunits are
transported out of the nucleoplasm through the
pore complex separately. They join only after
arrival in the cytosol and in the presence of
messenger RNA and transfer RNAs.
The nucleus is the site of mRNA
transcription
In addition to the production of rRNA and
tRNA, the nucleus is responsible for producing
messenger RNA. This process is called
transcription. Regions of the "active" genes in the
genome possess nucleotide sequences which are
called the promoter. This region is recognized by
RNA polymerase (a protein) as a site for binding to the DNA. The RNA polymerase slides down the length of the gene and catalyzes
the formation of a single-stranded RNA transcript. If this gene codes for a protein, the RNA transcript is called messenger RNA. The
processes of transcription and translation are depicted below.
The messenger RNA will be modified (introns removed, etc.) and shipped out of the nucleus through the pore complex.
In the cytosol, it will join to the small and large subunits of a ribosome and through the interaction of the ribosome with transfer
RNAs, a protein will be synthesized.
5. Plant Physiology
5
The ribosome is the site of protein translation
The ribosome subunits attach to the mRNA and together
slide along the RNA. As the subunits pass along the RNA, transfer
RNAs deliver amino acids as coded by the sequence of nucleotides
in the RNA. The ribosome proteins and ribozymes catalyze the
formation of the peptide bond between the amino acids to provide
the primary structure of the developing polypeptide.
Ribosomes can be found freely in the cytosol and
produce "soluble" proteins that are used locally in the cytosol.
Other ribosomes are associated with the endoplasmic reticulum.
These are attahed to mRNAs that code for a hydrophobic signal
sequence of 18-30 amino acids in the developing polypeptide.
These sequences bind to a signal recognition particle which
facilitates the attachment of the ribosome to the bilayer of the
endoplasmic reticulum and the penetration of the developing
polypeptide into the lumen of the ER. Once inside the lumen, the
polypeptide is processed in various ways to attach a particular
oligosaccharide to the polypeptide. This "label" facilitates the
sorting, packaging, and transport of the polypeptides to ensure their delivery to the correct intracellular or extracellular location.
The difference between rough ER and smooth ER is both structural and functional. Areas where the ER is associated with ribosomes
(rough ER) the main functions are translation of export proteins. Areas where the ER is not associated with ribosomes (smooth ER) carry
out synthesis of lipids and other membrane components. ER located near the nucleus is often rough ER; areas of the cell remote from the
nucleus are more likely to be smooth ER.
The endoplasmic reticulum processes and transports materials across the cell
The endoplasmic reticulum is a network of connected membrane sacs and tubules. Materials crossing into the lumen from the cytosol
are transported from one region of the cell to another. Precisely how the ER accomplishes the movements is apparently unknown.
Ultimately proteins that require packaging and export arrive through the network near an ending close to a Golgi apparatus.
Here the ER produces a vesicle that contains the protein and carries it across the cytosol toward the Golgi.
The Golgi (dictyosome) has two faces
The Golgi apparatus is very much like a specialized stack of ER; it is depicted. The layers of the stack which are closest to the ER
are called the cis face; the layers which are closer to the cell membrane are called the trans face. Each layer in the stack is called a
cisternum.
The cisternae of the cis face of the Golgi receive the vesicles from the ER with their contents. Within the cisternae, the
oligosaccharides at the end of the protein are modified again prior to export. Cisternae on the trans face either produce secretory vesicles
or disintegrate into large populations of export vesicles. In the latter case, new cisternae are produced on the cis face to replace those
breaking up on the trans face. Which of these two mechanisms, or some combination, is reality has not been determined precisely.
The secretion vesicles can migrate to the cell membrane and participate in exocytosis.
One type of vesicle that is critical for plants is a secretory vesicle that releases cell wall oligosaccharides and wall- enzymes by
exocytosis into the cell wall environment. Another vesicle, found in storage cells, are coated with clathrin and contain proteins and other
materials for digestion. These transport materials to special vacuoles for intracellular digestion. There are also means for lysosomes to
participate in this process.
6. Plant Physiology
6
The vacuole is a specialized and complex organelle
The vacuole is essentially a very large smooth vesicle, but this appearance is deceiving! From a membrane point of view the
vacuole is just a big bag, the tonoplast. The vacuole contents have little to offer visually, hence the name "little vacuum:"
Of course pigments are sometimes stored here, for example anthocyanins, and thus they were known even a century ago not to be
completely empty or just water. Indeed the vacuole is mostly water, as is the cytosol. This water supply of course serves to provide an
available supply of water, makes a large cell out of a small amount of metabolically expensive cytosol. The water in the vacuole helps
provide the turgor pressure for cell growth and tissue support. To keep water moving into the vacuole by osmosis, the vacuole
accumulates a range of solutes: minerals, ions, organic acids, polyphenols, sugars, enzymes, and a diverse array of relatively toxic
substances.
The solutes in the vacuole are toxic to herbivores
The solutes of the vacuole do more than provide an osmolarity to bring water into the cell. They do more than paint flowers red
and blue. The toxins are more than just stored away here. In fact animals eating plants get doses of these toxins.
Functionally the vacuole is a recycling center
But what is more important from the plant cell's point of view, are the enzymes located here. The enzyme array of
vacuoles is impressive. These include lipases, glycosidases, nucleases, and proteases: the range of enzymes needed to digest just about
any cellular component. These make the vacuole the plant equivalent of a lysosome in animal cell. In recent years we have learned much
about the recycling capacity of the vacuole. Materials are delivered into the vacuole, are digested into raw materials, and those are
returned to the cytosol for reuse. Obviously our thinking about vacuoles has changed greatly over the past century: from an empty bag of
water, to toxic waste storage pool, to component recycling center.
Plants have mitochondria for respiration
Yes, plants have mitochondria. Many people do not realize that plants have mitochondria and carry out respiration. This
misconception needs to be repaired immediately. Surely most plants obtain most of their energy through photosynthesis, but this does not
mean that respiration is unimportant. Quite the opposite is true! All living plant cells survive the night time and cloudy days by
respiration, most of which takes place in the cell's mitochondria. Consider root cells...under the soil there is no light penetrating to drive
photosynthesis; its entire energy needs must be satisfied by respiration! Indeed all living cells of plants (except for sieve tube members
which use companion cell contributions instead) have mitochondria that assist with respiration.
The mitochondrion is probably derived from an
endosymbiont
The mitochondrion consists of a smooth outer membrane and a
much larger inner membrane; the inner membrane is about 70% protein
and is highly convoluted to fit inside the outer membrane. The space
between the two membranes is called the intermembrane space. The fluid
inside the inner membrane is called the matrix. This matrix is believed to
be the cytosol of an ancient prokaryotic endosymbiont. It contains
prokaryotic style 70S ribosomes and its own genome, the chondriome, in
the form of a single, circular DNA molecule which is not associated with
histones. The genes in this DNA code for some of the critical enzymes
and electron transfer proteins needed for the Kreb's cycle and the electron
transfer chain which are translated on mitochondrial ribosomes. The key
word here is "some;" indeed many of the components in the
mitochondrion are coded by DNA genes which are now housed and
transcribed in the eukaryotic nucleus and are translated on cytosolic ribosomes. Mitochondria replicate their naked chromosome in a
process similar to fission in baceria; they divide by a process similar to cytokinesis by furrowing.
Mitochondria host the Kreb's cycle, electron transfer chain, and oxidative phosphorylation
7. Plant Physiology
7
Indeed plant mitochondria are not much different from those in animals. While they may be more round than oval in shape, plant
mitochondria carry out the Kreb's cycle in the matrix and operate the electron transfer chain in the infolded cristae of the inner membrane.
The electron transfer chain also pumps protons (H+) into the intermembrane space. The pumping and diffusional gradient between the
space and the matrix represents a conservation of energy from respiration. As protons leak back into the matrix from the intermembrane
space through an ATP synthase membrane protein complex, this potential energy is trapped in the terminal phosphate bond of ATP.
These structural and functional ideas are partially shown here.
Chloroplasts are also likely to be derived endosymbionts
Chloroplasts are likely derived from endosymbiotic
cyanobacteria. They have two smooth membranes (inner and outer)
surrounding the stroma. The membranes are likely derived from the
cell membrane of the endosymbiont and the stroma is its cytosol.
The membranes are made of mostly of glycosylglycerides rather
than phospholipids.
The stroma contains 70S ribosomes, developing starch
grains, oil bodies, and the naked, circular DNA chromosomes. Also
present in the stroma is an endomembrane system called thylakoid
lamellae. Some of the lamellae are stacked as grana; other lamellae,
called stroma lamellae, interconnect the grana. The chloroplast
genome (the plastome) codes for many, but not all, of the proteins
required for photosynthesis. The transcripted RNA is translated by
the 70S (prokaryotic style) ribosomes.
Other proteins are now coded in the nucleus and translated
in the cytosol; these notably include the large subunit of RuBP
carboxylase/oxygenase. The chloroplast genome is 145 kbp
compared to 200 kbp in the mitochondrion and together these
genomes represent perhaps 1/4 of the genes in the cell.
The chloroplast carries out photosynthesis
The stroma of the chloroplast hosts enzymes which carries out the
Calvin cycle reactions to convert carbon dioxide to carbohydrate. The
endomembrane system hosts the electron transfer proteins and pigments for
the light reactions which split water and produce oxygen gas. These two
reaction systems are coupled; each reaction system needs some products of
the other.
The electron transfer proteins of the light reactions pump protons into
the thylakoid lamellae; the protons pass back through ATP synthases of the
thylakoid membrane on their way back to the stroma; this results in ATP
synthesis as in mitochondria. The
ATP is used by the Calvin Cycle. Thus both systems require
environmental light and carbon dioxide. There is no such thing as a "dark
reaction." The carbohydrate product of photosynthesis can be accumulated in
the chloroplast in the form of a developing starch grain or as lipid bodies.
Microbodies
Microbodies were the original name for small single
membrane bound organelles. In plants these have been renamed
peroxisomes and glyoxysomes. The peroxisome is shown with its
crystalline matrix of catalase below. This organelle mostly degrades
glycolate, a 2C acid produced in chloroplasts as the result of
RuBisCO combining with oxygen rather than carbon dioxide, in the
process of photorespiration. The glycolate is transferred into the
peroxisome. In degrading glycolate, oxygen is consumed and
peroxide (H2O2) is produced. This toxic material is enzymatically
degraded to water and oxygen by the enzyme catalase. This critical
enzyme may comprise 40% of the protein in the peroxisome; little
wonder catalase appears in crystalline form in peroxisomes.
Mitochondria are a third partner in photorespiration.
Oleosomes store oils
The partner for glyoxysomes are oleosomes. These
organelles are spherical and are bounded by a phospholipid
monolayer. The hydrophilic polar phosphate "head" of the phosopholipid faces the cytosol and the hydrophobic fatty acid "tails" face the
interior of the oleosome which is loaded with hydrophobic oils from the ER. There are peripheral proteins on the hydrophilic side of the
monolayer. These proteins include oleosins which may help attach the enzyme lipase to this monolayer to initiate fat digestion for the
glyoxylate cycle.
8. Plant Physiology
8
Glyoxysomes carry out the glyoxylate cycle
Glyoxysomes are abundant in oil storage tissues. This microbody
ontains the enzymes needed to degrade fatty acids into 4C acids to drive
mitochondrial respiration and reverse glycolysis to make sugars
(gluconeogenesis) for growth and development. This pathway is called
the glyoxylate cycle. This organelle is active as the oils are removed from
storage and put into use or exported for transport in phloem. You will
notice how, just like photorespiration, this pathway involves three
organelles and the enzymes in the cytosol to complete the conversion of
oils back into carbohydrates.