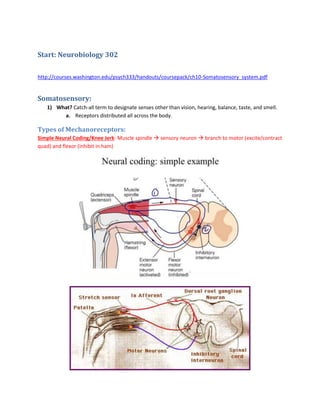
2. http://www.ifc.unam.mx/Brain/reflex.htm
1) Kneejerk Reflex: Tapping the knee pulls the tendons of quadriceps.
a. When this muscle stretches, information in the form of APs goes down (1) through
the sensory neuron. This is because the stretch-sensitive receptors (spindles) are
excited. This spindle is innervated by the 1a afferent fibers, which lead to the spinal
cord.
b. Sensory neurons synapse with extensor and flexor motor neurons in the spinal
cord.
i. Extensor/Excitatory: Contracts the quadriceps, the muscle that was
stretched.
ii. Flexor/Inhibitory: Inhibits flexor muscles that would contract the
hamstring.
Frequency Coding:
Linear relationship when transducing from intensity to frequency.
3. Concept of Receptive Field:
1) Definition: Location or sensory space in which a stimulus will elicit a response from the
neuron.
4. a. Example: Any part of skin that, when stimulated, will elicit a response from that
receptor.
2) Each sensory system has its own definition of receptive field.
Somatosensory Submodalities:
1) Definition: Specific sensations encoded by the somatosensory system.
a. Modality simply means stimulus type.
5. 1) The submodalities are mediated by dorsal root ganglion cells, which are where the cell
bodies are located.
a. These dorsal root ganglion cells are composed of similar-type cells that act as the
sensory neurons for the somatosensory system.
2) These cells are bipolar:
a. One axon goes to the periphery (skin, muscle, joint capsules, etc.) with specialized
receptors for specific types of stimuli.
i. This axon is ‘dendrite-like’ in that it receives information from the outside
world, but it isn’t actually a dendrite.
ii. Note that there’s a freely branching ending at the end of the periphery
axon.
b. Second axon goes to the CNS through the dorsal root of the spinal nerve.
3) Both of these axons are myelinated for really fast conduction.
6. 4/1: Mechanoreceptors in the Skin:
Tactile sensations in the human hand arise from
four kinds of mechanoreceptors: Meissner corpuscles (RA1 - low frequency/amplitude (light) skin motion
– lateral hand motion across objects … makes sense because upper layer/light touch)
Merkel cells (SA1 – edges and points – slowly adapting so good for detecting constant pressure), Pacinian
corpuscles (vibration – rapidly adapting good at distinguishing frequency changes – on off on off), and
Rufini endings (SA2 – stretching is a constant pressure, good for slowly adapting)
Summary:
Corpuscles/Rapidly Adapting (1 is epidermal, 2 is deeper, dermal).
Misses’ go first.
7. 1) Hairy Skin: Nerve terminals are wrapped around the base of each hair.
a. Hair motion is encoded based on direction and intensity.
Our focus is on the Glabrous Skin (Smooth Skin):
There are several different types of mechanoreceptors (touch receptors) in the skin.
8. 1) Unmyelinated Nerve Endings (1): Unmyelinated nerve endings can go to many different
places and are involved in chemical inputs and pain.
2) Epidermis:
a. Meissner Corpuscle (RA1): Unmyelinated nerve endings can attach to the
Meissner Corpuscle.
b. Merkel Disc Receptor (SA1): “ “ can attach to this as well.
3) Dermis:
a. Unmyelinated endings can also attach to the Ruffini Endings (SA2) or Pacinian
Corpuscle (RA2).
4) Recall that the other side of the myelinated axon goes over to the CNS (other side of the
peripheral nerve bundle).
Meissner and Merkel:
9. 1) Recall these are the ones located in the epidermis.
2) Meissner Corpuscle (RA1): This is the receptor organ a globular, fluid-filled structure of
flattened cells with nerves between the layers.
a. Rapidly Adapting: If we record from the nerve and stimulate as a ramp function,
we see that the receptors soon stop generating APs.
3) Merkel Cell (SA1):
a. Rigid structures that transfer strain from surface to nerve ending.
b. Slowly Adapting: If we record from this cell, a constant stimulus will not lead to
adaptation. It will thus continue to respond to pressure for as long as it is sustained
(within a reasonable time frame).
c. Use: Edges/corners/points.
Pacinian Corpuscle and Ruffini:
1) These are located in the dermis, deeper layer.
2) Pacinian Corpuscle (RA2): Large, onion-like capsule.
a. Rapidly adapting, so we stop firing as soon as the indentation is stationary.
3) Ruffini Endings (SA2): Primarily excited by stretching – can tell how much stretching is
on the hand after holding an object, for example.
Concept of Best Frequency:
1) What? Idea that each receptor is most sensitive to stimulation at its natural or best
frequency.
2) Merkel Cell vs. Meissner Corpuscle:
10. a. When we push on the corpuscle, the structure itself moves in response, but we
adapt quickly.
b. By the frequency encoded, we can determine if something is moving or sustained.
c. Example: Bug moving around on your finger. This is encoded by the Corpuscle (RA)
as a rapidly adapting response that soon turns off. It detects change in position of
the bug.
i. On the other hand, the Merkel (SA) will detect that the bug is still there
(motionless). Remember it is slowly adapting, so it can detect deformation of
the skin (pressure).
Receptive Fields of Mechanoreceptors:
1) What? Individual mechanoreceptors convey information to a limited area of skin called
the receptive field.
2) How? Mapped by recording from median nerve and stimulating hand with different
sizes/strengths of stimuli.
Superficial vs. Deep Layers Comparison:
11. 1) Superficial (Type 1): Relatively small receptive field consisting of small spots.
a. Graph C shows a zoomed in version of the receptive field of a single fiber/corpuscle.
2) Deeper (Type 2): Innervate deeper layers
a. Activated when you press deeper on the skin.
b. Differences: Receptive field has one hot spot whose sensitivity is greatest (located
directly above receptor) with an overall wider arc surrounding this hot spot.
i. Possibility for overlap of inputs with other nearby Pacinian corpuscles.
12. Rapidly Adapting Pacinian Corpuscle:
1) Corpuscle: Made of connective tissue; the unmyelinated part of the RA2 fiber is located
inside.
2) Response: when sustained input/pressure is applied, the RA2 fiber fires a burst at the
start and end of stimulation
a. Why? At the beginning, the connective tissue deforms and then quickly adapts
to the stimulus. So, the underlying nerve stops feeling the pressure and stops
firing. When we take this stimulus off, we redisturb the tissue and underlying
nerve, making it fire at the end.
i. Contrast to sinusoidal firing (vibration), which causes a continuous
firing – no adaptation because not constant (vibration-like).
Experiments on Pacinian:
13. 1) This shows that the rapidly adaptation of the Pacinian is wholly due to the physical
surroundings of the nerve endings: the connective tissue surrounding the nerve fiber is
what causes the rapid adaptation.
a. We have a sustained receptor potential when the connective is removed (no
adaptation).
1) Y-Axis: Amplitude of the vibration.
2) Left Graph: Shows that the best frequency is around 200 Hz for the RA2 fiber.
a. We only need a small amplitude vibration for a person to notice something
happening.
14. b. At lower frequencies, say 10 Hz, it requires more amplitude to feel something going
on.
3) Right Graph: What a person actually perceives in a situation is exactly the same as taking
all of the mechanoreceptors and looking at the most sensitive receptor at each frequency.
The ‘real’ frequency matches the most sensitive frequency.
Two-Point Threshold:
1) What? Measures the minimum distance at which two stimuli are resolved as distinct.
a. We use two prongs to touch various parts of the body.
b. For example, we would feel two distinct prongs if we touched the fingers (smaller
threshold) vs. the back (would feel only one).
c. The fingers, lips, soles of feet are most sensitive (receptive field smallest).
i. More receptors in smaller receptive field.
15. ii. More receptors means better ability to discriminate two points.
2) A place on the body that is more sensitive has a smaller threshold for distinguishing two
points. At any distance above that threshold, we would distinguish two points. Anything
smaller than that we would only smush it to one point.
4/2: KCNQ4 and Somatosensory Continued
1) SA1 Merkel: Most distinct and fine. Nerve fires only when moving over the dots and is
silent in the spaces (no stimulation).
a. Receptive field smaller than the size of dots each dot stimulates a new set of
SA1 fibers.
2) RA1 Meissner Corpuscle: not as good at distinguishing because of larger receptive
fields.
a. Function: Can tell whether something is present or not present.
3) RA2/Pacinian Corpuscle: Good at detecting vibrations, although there is no
discrimination – receptive fields too large.
4) SA2/Ruffini: Receptive fields also too big.
KCNQ4:
1) There are many different types of K channels – we’re going to focus on the KCNQ4.
2) Normally, the resting K channels are important because they set resting conductances for
the neuron.
16. 3) The KCNQ4 regulates cell excitability because it modulates membrane potential in cells
where it’s expressed.
The M Channel (blocked by Muscarine – Ach agonist)
1) Ionotropic Ach channel (fast EPSP) and then we get a slow EPSP (Ach activated a
metabotropic receptor second messenger system to close M-type K channel).
2) If we close these M-type K channels, we make the cell more excitable because we are taking
away the resting channels (threshold more negative).
a. Graph C shows this.
17. 1) Graphs plot value of current vs. each held voltage.
a. The I-V curve is shifted extremely negative.
b. The M-current turns on at -50 mV (resting potential of most cells).
c. This M current sets excitability of the cells. It is open near rest.
1) Block M current lower resting conductance, so the cell becomes more excitable.
18. Recall: At hair follicle mechanoreceptor, we deform free nerve endings (unmyelinated), physically
changing membrane which opens up ion channels, depolarizing and making APs fire.
KCNQ4 Channels in Mouse: NF200 stain axons, KCNQ4 stains for itself.
1) Hair follicle: Free unmyelinated endings wrap around the follicle.
a. Coexpressed KCNQ4 and axons in the unmyelinated endings.
b. Coexpressed in the corpuscle or in the hair follicles.
19. 1) Apply ramp of indentation to the messiner corpuscle:
2) Wild Type vs. Mutant: In mutant, we take out K channel more excitable
hyperexcitable mutant cell doesn’t adapt as well as the wild type.
1) Humans also have the KCNQ4 expressed in the hair follicle. However, some people do not
have this expressed – they are deaf.
2) In a) we apply a frequency of stimulus that continually increases. We ask when they feel the
stimulus.
3) KCNQ4 is expressed around hair follicle and Messiner – not found in Pacinian.
a. What does this do?
i. Frequencies felt by Pacinian are not changed – no KCNQ4
ii. Frequencies in hair follicle/messiner – the individuals without KCNQ4 are
better able to notice stimulus – lower amplitude threshold for a given
frequency (more sensitive and excitable/more negative threshold).
iii. More sensitive (require smaller amplitude stimulus) to low frequency
events.
Spinal Cord:
Question: What happens to the signals after they hit the mechanoreceptor?
20. 1) Conduction velocities correspond to different nerves in a bundle.
2) X-Axis is conduction velocity.
a. C fibers conduct slowly because they are unmyelinated – involved in pain.
3) In the others, we have a 6x relationship between diameter and velocity (myelinated).
a. Cold and hot temperature/pain pathway (A delta)
4) Mechnoreceptor (A beta) – 12 72 diameter/velocity. We’ve been talking about these –
the peripheral mechanoreceptors.
a. Found in cutaneous nerves.
5) A alpha: large, muscle spindle
Spinal Anatomy:
21. 1) 31 pairs of spinal nerves
2) Throacic (trunk)
3) Lumbar, sacral (Legs)
4) Spinal nerves are paired: one on left and one right.
5) Each dorsal root innervates a segment of skin of the animal – segmented dermatome.
a. Shingles dermatome.
22. 1) Mechanoreceptor signals come in the spinal cord at the dorsal root ganglion and diverge
in two ways:
a. Sensory Neuron Motor Neuron: Tells the muscle that sent the signal to move.
b. Brain Stem: Higher up signal processing.
4/4: Anatomy of Spinal Cord/CNS:
This figure shows how somatosensory inputs are translated up to the brain stem.
1) Axons of motor neurons are preprogrammed during development to seek out the ventral exit
point (ventral roots) periphery to target muscles.
2) Dorsal Root contains the central and periphery projections.
a. Once axons from periphery enter dorsal root branch to motor neurons (where exactly
depends on modality of stimulus) OR
b. Can branch to the ascending (labeled 2a) go up dorsal columns to the brain stem.
3) The modality of stimulus is encoded in the mechanoreceptor itself. It is then “faithfully
transmitted” from the periphery into the CNS through axons.
a. The axons are ‘passive railways’ for the signals to pass through after encoded by the
receptor.
Anatomy of Spinal Cord:
1) The Honda-shaped structure that I outlined has dark blotches that are cell bodies (grey
matter).
2) The surrounding white matter consists of axons (myelinated axons appear white).
3) The upper splotches are interneurons that receive sensory information and project the
signal to the brain stem.
23.
24. 1) Grey: White Matter Ratio Increases ROSTRAL CAUDALLY: There is more white matter
(axon-containing parts) in the cervical because the axons that start in the lumbosacral
(gracilus fasciulus) continue up past into the cervical, taking up space in the white matter +
the new white matter from the cunenate fasciulus (cervical).
a. Lumbosacral: Just axons from there!
b. Cervical: Axons from lumbosacral AND axons from cervical area means more white
matter (outside part) and less grey matter.
25. A) Receptors land in different layers of the spinal cord (I- X) – each class of DRG finds its way to a
specific portion of motor neurons.
26. 1) Cervical/Lumbar vs. Midthoracic: There is more grey matter in the cervical portion because
that’s where the limbs (specifically, arms) are located than in the thoracic (trunk).
2) Consequently, we get more motor neurons located in the cervical more grey matter.
27. Dorsal Column Medial Lemniscal Pathway:
1) Move from dorsal part of the spinal cord to the medial-lemniscal.
2) Pathway: Finger with mechanoreceptor (input) dorsal root ganglion one axon
branches to the motor neurons, one axon goes to dorsal (brainstem) medulla and cross
over (decusses to the other side) to become the medial lemniscus axons go to
thalamus.
What is the Thalamus? Also called ventral posterior nucleus
1) Thalamus is the waystation nucleus for our senses.
a. Functions to deliver signals to different parts of the cortex depending on the type of
stimulus (pain, visual, mechanosensory, etc.)
b. Codes for type of sensory stimuli, where it was in our body, and what type of response is
required.
28. __
Next Step: Reassembly at the Cortex.
2) Sensory information relayed to the cortex by the waystation thalamus.
a. Various types of info are laid out and segregated and joined again in cortex.
b. Types of sensory information are sorted out by the thalamus and relayed/put back
together in somatosensory cortex (S1).
i. Inputs from thalamus go to S1, A1 (auditory), V1 (Visual)
Anatomy of the Brain: Four Regions
1) Occipital (visual input and processing)
2) Temporal Lobe (auditory, speech)
3) Parietal Lobe (Somatosensory inputs land here)
4) Frontal Lobe (Plan movement in front portion, execute movement as you move back).
Anatomy of the Cortex:
29. Three Cortexes: Primary (sensory), Motor, and Association Cortex
1) The cortex is like a cloak that lies on the very top part of the brain.
a. The cortex consists of grey matter (cell bodies).
b. Everything else is white matter (axons).
2) All the units of the cortex are linked together via axons. Axons can travel long distances
from the cortex or short distances.
30. 1) The Cortex is Striated: This physical striation reflects the fact that sensory information is
sent by thalamus to specific part of cortex. The information lands in different
layers/striations.
2) The Cortex has Six Layers (Applies to all three types of cortex):
a. I is very small in adulthood (outer layer of the brain)
b. Primary Sensory Cortex:
i. Layer Four is the input layer. The cells in layer four are accepting input and
project to the other layers of cortex. The layer is broken up into sublayers
(A, B, C).
1. So, it is the largest layer because it takes in a lot of information.
ii. The other layers are projection neurons that send information to other
parts of the brain.
c. Primary Motor Cortex:
i. Very small IV layer, but V and VI have huge cells (Betz cells - motor
neurons) that might make a twitch in the finger if we stimulate this area of
the cortex.
31. 1) We get lighter moving from primary sensory cortex to association cortex because
there are fewer cell bodies in that area. Recall layer four in sensory had a lot of stuff,
association has less (no stripe).
a. Notice the really dark portion in the sensory – that’s layer IV.
b. This is conserved throughout the visual, somatosensory, and auditory system.
The layer IV is always denser because that’s where the axons input.
32. Somatosensory in Detail:
1) Note the layers – that is where we would see the layers we talked about earlier.
2) Central sulcus divides motor and somatosensory cortex.
3) Inputs coming in from dorsal column are laid out in the cortex. They are arranged in regions (3,
1, 2) based on where it comes from in the body and its modality
a. Note 3a and 3b are wrapped around the sulcus (dips inward).
33. 4/7: Somatosensory Details
Note: This figure shows the area of the cortex dedicated only to one middle finger (see below
figure).
1) Recall: Each mechanoreceptor detects a different modality (type of touch – vibration,
pressure, etc.)
a. We separate the inputs in the thalamus by modality and location of input and then
reassemble this information in the cortex.
2) Region 5: “Active Touching” is responsible for “actively” trying to figure out what
something is.
3) Different mechanoreceptors are sent to different regions of the cortex.
4) Convergence of Information to Area 2: Neurons within 3a, 3b, 1 send their axons to each
other through the white matter (for example, see an arrow that goes from 3a to 2).
a. All inputs from muscle spindles, sa1, ra1, ra2 all end up at area 2.
5) Figure B/Receptive Field Size: The receptive fields get larger because we’re adding more
information from more receptive fields – we are summating the receptive fields.
a. After combining the info in region 2, the cortex can identify can determine
attributes of the object.
34. Close-up of digit 3 AKA middle finger.
1) The inputs initially land in layer IV and are then sent to different regions. The axons in layer
IV then synapses with projection neurons, which are then sent to section 2 in the cortex
(see last figure).
2) Note that the fingers are organized right next to each other on the cortex (zoom in figure A).
a. The fingers are represented in both section 3b and section 1 – they are both input
layers.
35. 1) Different parts of the body is partially represented in many different places.
2) Cortical Amplification: More cortex is dedicated to the fingers vs. the torso because there
are many more mechanoreceptors in the fingers vs. torso.
1) The somatosensory map represents how much cortex is dedicated to each part of the body.
2) This is composed of a cross section of the somatosensory cortex.
3) More mechanoreceptors and smaller receptive field in the larger “pieces”
a. Smaller receptive field means more densely packed axons, meaning more axons
going to the cortex.
36. 1) Example: Playing piano. We can compare before vs. after training. The digit that got the
most training has more representation in the cortex.
a. The receptive fields are also smaller.
1) Example: Removing Limb. We get phantom responses. This is caused by rearrangement
of cortical circuits.
a. Adjacent fibers expand. Now, this place that used to be innervated by axon fibers
from the hand are innervated by other parts of the skin.
b. Brain interprets activity from face/upper arm as from the amputated limb.
37. Comparative Physiology:
1) Vibrissae: Nocturnal rodents use this mechanoreceptor for whisking – tells the rodents
what kind of stuff they’re walking in.
a. Each vibrissae represented on somatosensory – vibrissae highly represented in S1
b. Each hair has innervating nerve endings that go through the typical pathway
through thalamus and cortex.
2) Staining in the second figure shows where oxygen is being used. We can see the barrels of
vibrissae, all of which land in layer IV.
The Star-Nosed Mole:
1) The nose has a star on it! There are mechanoreceptors on the rays.
2) Receptive fields near the middle (1 and 11) are the most sensitive – allows mole to
determine what it has is food and if so, it will eat.
3) The other rays consist of mechanoreceptors (free nerve endings) with Merkel cells below,
then something resembling a Pacinian Corpuscle below that.
a. Myelinated nerves go through the typical pathway to the thalamus then
somatosensory cortex.
39. 4/8: Pain Pathways
1) Pain is mediated by nociceptors, made of free nerve endings of primary sensory neurons.
2) Largest axons are the A alpha and A beta, followed by A delta then C fibers.
3) Result: When we experience pain, we have two types of pain.
a. The first is the sharp pain. This is transmitted by the A deltas.
i. The A alpha/beta are just detecting change in skin, no pain involvement.
b. The second is the more prolonged, burning pain.
i. Mediated by the C fibers.
40. 1) Minor shock: A Alpha activated, fastest conduction velocity but barely perceived.
2) Medium Shock: A alpha and A beta (mechanoreceptors recruited).
3) Strong Shock:: A delta added on (sharp pain)
4) Strongest: C fibers activate later, causing the burning pain.
41. 1) TRP (Transient receptor Potential) ion channels convert noxious stimuli energy into a
depolarizing electrical potential.
a. These are expressed by nociceptive neurons. They are located at the free nerve endings
of the C fibers.
2) There is a wide variety of TRP channels, which mediate different sensations (temperatures and
chemicals).
a. TRP1 responds to cold, TRPV2 responds to hot.
b. Some also respond to chemicals.
3) Determined by seeing the best response of the channel through whole-cell recording and
changing temperatures.
42. Pain Pathway:
1) Synapse onto projection neurons (substantia gelatinosa, SG). These neurons cross to the other
side of the spinal cord (desucces) and then go straight up to the brain through thalamus.
2) This pathway for pain is different and is called the spinothalamic pathway.
a. After synapsing with the SG neurons (projection neurons), the axon goes straight up to
the thalamus.
43.
44. Pathway for Mechanosensory vs. Pain:
1) Mechanosensory/Dorsal Column-Medial Lemniscal: Hits receptor DRG birfurcates
one axon goes through the dorsal column lemniscal pathway after it decussates in
medulla.
2) Pain/Antereolateral: C fibers from nociceptor go through a different DRG cell and synapse
onto a projection neuron (substantia gelatinosa) in the dorsal horn. This then
decussates and goes up the antereolateral pathway.
3) Note: See that mechanosensory input decussates at the medulla. In contrast, pain inputs
decussate in the dorsal spinal cord.
45.
46. Gate Theory of Pain:
+ means excitatory, - means inhibitory.
1) Two different receptors: one mediates pain, another mediates mechanostimulation.
2) C fiber (pain) synapses with the projection neuron.
3) C fibers also innervate an inhibitory neuron.
4) Follow the red pathway for C pain fibers.
a. C fibers synapse and excite the pain projection neuron (more pain).
b. But, we can also cause pain by inhibiting the inhibitory interneuron (double negative, so
it excites the pain neuron).
5) There are also inputs from the regular mechanosensory axons which synapse onto inhibitory
interneurons and projection neurons.
a. Can excite projection neuron. This happens when we stimulate the skin a lot, causing
pain.
b. “Rubbing to Decrease Pain:” Can also excite the black interneuron to inhibit the pain
projection neuron. This causes less pain.
6) The Gate Theory: Projection (pain) neurons receive mechanosensory and nociceptor
information.
a. Inhibitory neurons inverts signal, giving us the “rubbing to decrease” phenomena.
47. 7) A high frequency of stimulation to the mechanoreceptor can cause pain. There is a greater
“factor” of stimulation to the pain projection neuron vs. the inhibitory interneuron. So, we can
cause pain if we rub something too hard.
Control of Pain:
1) There are two pathways for the control of pain.
a. Red Line: Natural opioids release and activate serotonin, inhibiting nociceptive neurons
(decrease pain).
49. This is a cross section of the SPINAL CORD before we reach the medulla.
1) Medial Lemniscus (Top): Crosses the midline in the medulla. The somatotopic
representation gets inverted.
a. Displays head medially, sacrum laterally, hands and feet ventrally.
2) Spinothalamic Pathway/Pain: Cross in spinal cord.
a. Fibers originating in lumbar and sacral are located laterally, while those from
the cervical spine (C) are positioned medially.
3) Since the somatosensory pathway doesn’t cross until the brain (medulla) and the pain
pathway crosses in the spinal cord itself, we get inversion.
4/9: Visual System
Nice powerpoint follow along
http://www.ic.ucsc.edu/~bruceb/psyc123/Vision123.html.pdf
50. Basic Anatomy
General Anatomy of Retina:
1) Thin black layer shown is the pigmented epithelium.
a. Functions to 1) absorb light of extra light bouncing around retina 2)
regenerates/maintain photoreceptors.
2) Retina composed of a thin sheet of neurons. They and the optic nerve are part of the CNS.
3) At the fovea, neurons are shifted aside so light goes directly to the photoreceptors.
a. Many photoreceptors are packed in the fovea. These are all composed of cone
photoreceptors, designed to see things with high accuracy.
51. 1) Light goes through layers of cells before it impinges on photoreceptors.
a. Cells are pushed aside at the fovea, though (not shown here).
2) RPE composes the outer layer and we move inwardly from there.
3) Outer nuclear layer: Contains the nuclei of the photoreceptor cells.
a. These cells transduce the signals – signals sent the opposite direction that the
light is coming in.
4) Inner Nuclear Layer: photoreceptors synapse onto bipolar cells.
a. Bipolar because they spread in two different directions.
b. This layer also contains horizontal and amacrine cells which summate
convergent signals (will talk about in detail later).
5) Not all of these cells fire action potentials.
a. The only cells that spike are the ganglion layer cells (output cells to the optic
nerve and cortex).
b. We need a spiking output at the top. The different types of lower cells merely
mediate the different types of the light that comes in.
52. 1) Light impinges on the photoreceptive pigments at the bottom, then sensory information
goes back up.
53. 1) Photoreceptors are on top.
2) Outer nuclear layer has the cell bodies of the photoreceptors.
3) Inner nuclear layer contains cell bodies of horizontal cells and amacrine cells.
4) Ganglion layer sends out information to the optic nerve.
5) Conduction in visual system is slow because we go through both second messenger
pathway and several synapses before even hitting the CNS.
6) The different types of bipolar cells extract specific types of information from visual field.
7) Rods and cones synapse in different locations in the retina.
54. 1) The blind spot is located where the optic nerve comes in (NOT the place the light is
pointing right now).
2) All the axons in the retina have to go to the optic nerve to get into the CNS.
a. There are no receptors over the optic nerve, so we have a blind spot.
55. 1) Recall the fovea, the point of sharpest focus. The density of photoreceptors, bipolar cells,
and ganglion cells is highest here.
2) Fovea/Cones: In the fovea, there is a 1:1 relationship between cone: bipolar cell, meaning
we have much higher acuity (vision is sharpest). Information about the object is
preserved or even enhanced.
56. 3) Periphery/Rods: As we move away from the fovea, we get more convergence. If many
photoreceptors converge into one bipolar cell, we have lower acuity. This is because more
convergence means pixels get bigger, less fine.
a. Also amplifies signal as well by adding together activity from many photoreceptors.
b. See in the dark, but acuity is not as good, though.
4) Note the relative amount of cones vs. rodes in the fovea - fovea basically made of all cones.
Types of Photoreceptors:
1) Split into rods and cones.
2) The cone has a higher membrane capacitance because there’s more surface area due to
folds.
3) Outer segments contain the photoreceptors.
59. 1) Rhodopsin is the visual pigment in rod cells. It is made of a protein component and light-
absorbing component.
a. Protein component: Opsin.
b. Light-Absorbing Component: Retinal.
2) Opsin, the protein, wraps seven times across the membrane of the rod.
3) Retinal absorbs light changes conformation (11-cis all-trans) expanding and allowing the
protein on the membrane to uncoil. This activates second messenger system.
Detailing the Transduction Pathway
1) This stuff was worked out in rods because there is a high density of pigments there.
2) In the dark, we have cGMP gated channels open because cGMP is high, allowing Na+ and
Ca2+ to come in the cell.
a. Steady influx of Na+ maintains cell at depolarized -40 mV.
i. Result: Constant release of glutamate
3) In the light, rhodopsin is excited by absorption of photons, activating transducin
activate phosphodiesterase dropping cGMP levels closing cGMP-gated channels.
a. The alpha and beta components of transducin (T) separate. The alpha component
activates PDE. PDE breaks down cGMP into GMP. Since cGMP levels are lower, the
channels close, hyperpolarizing the cell.
b. cGMP channels are dependent on cGMP. The more cGMP we have, the more
channels open.
60. c. In this scenario in the light, less Ca and Na come into the cell, hyperpolarizing the
cell.
4/11: Visual System Continued
1) All of this transduction is happening in the outer segment.
2) When the cGMP channels open and we depolarize the cell, we release glutamate.
3) Dark: Since the channels are open, we are depolarized. Glutamate is released at high
concentrations.
4) Light: Channels close, hyperpolarization, less glutamate release
a. Figure C: More light intensity = more hyperpolarization because more channels
close.
61. In RODS!
1) Note all the amplification that occurs after one photon of light is absorbed by rhodopsin!
2) One photon thus affects membrane potential quite a bit, sending a signal to the brain that
light is in the visual field.
3) This is not a rapid response compared to mechanoreceptors because it has to go through
this long pathway!
1) This is the reason why it’s very hard seeing longer wavelengths in the dark (like red).
Rhodopsin can’t pick up those wavelengths.
2) Notice the correlation between the wavelengths for human perception vs. what rhodopsin
absorbs.
62. a. Proves that we’re basically using rods in the dark because no cones are involved
here.
63. 1) Since there is processing already in the retina, unlike the mechanoreceptors, the receptive
field is defined differently.
2) There is a donut of cells surrounding the pinkish photoreceptors in the middle. These
surrounding cells are also able to change activity when they synapse to the same bipolar cell.
3) We essentially get a larger receptive field produced as we move “further down” the pathway of
transduction.
i. This is because we have convergence of rods onto bipolar/ganglion cells, “summing”
their receptive fields as we move downstream.
4) Fovea vs. periphery (top part of figure):
d. Fovea has much smaller (blue dots) receptive fields because fovea has mainly
cones, which are 1:1 ratio between cones and bipolar.
e. Convergence of photoreceptors in the periphery, little in the fovea.
64. Bipolar Cells:
1) Function: Receive input from photoreceptors.
2) Note that the axons/synapses land in different layers (off vs. on plexiform layers),
meaning they have different functions as well.
65. What do mean by off vs. on?
1) Bipolar cells are the pathway from photoreceptors to ganglion cells.
a. Sign is conserved between the bipolar and ganglion cell.
66. b. Ganglion cells are the ones that actually fire.
2) Divergence: Cone synapses onto many bipolar cells.
a. Function: Allows for parallel processing.
b. Note that one bipolar cell is off-center, another is on-center.
Concept of On vs. Off Center:
1) We examine the output response from these neurons.
2) Off-Center: Depolarization in the dark.
a. Dark: Opposite of what’s shown in the figure. Glutamate is released from the cone
cell and activates ionotropic AMPA receptors, causing depolarization.
i. Same thing happens in the next step to the ganglion cells. This means we
have a constitutive inward current (depolarization).
b. Light: Opposite as in the dark. We remove the constitutive inward current,
hyperpolarization meaning less glutamate is released. The ganglion fires less.
This is what we mean by off. When we shine light on these cells, they stop firing.
3) On-Center: Fires in response to light.
a. These cells express a different metabotropic glutamate receptor.
b. Dark: When glutamate is released, we actually hyperpolarize the cell by opening
K channels.
i. So, we decrease firing rates in the dark.
c. Light: Opposite. We depolarize by closing K channels. More glutamate is released
because of the depolarization, increasing firing rate of ganglion cells.
The last point shows the significance of the parallel processing.
67. Within the receptive field, we can distinguish between center and surround regions, where light causes
opposite responses. The surround region is antagonistic to the center region.
1) In the indirect pathway, we factor in the horizontal cells.
a. This indirect pathway is termed the center surround. It uses GABA (see next
slide).
2) We have overlapping receptive fields – we can consider each receptor as both a center
and surround.
3) Center and surround are always antagonistic to one another. The light in the surround
hyperpolarizes and thus antagonizes.
68. 1) Horizontal cells (H) release GABA and inhibit.
2) On the left side, GABA release is constitutive from the horizontal cell.
3) When we shine light on the direct, off-center pathway cone (C), C hyperpolarizes less
glutamate is released bipolar cell (B) hyperpolarizes.
69. 1) Light to the surround.
2) When we shine light on the surround, we have hyperpolarization less glutamate is
released from the left cell. This means less GABA is released by the horizontal cell (less
constitutive hyperpolarizing current).
a. Less inhibition means more depolarization at cell C and more glutamate released
at C, meaning depolarization in the bipolar cell (B).
3) This is antagonistic to what happened when the center itself was illuminated.
a. See last slide. It hyperpolarized vs. now it’s depolarized.
For ON-Cell, we still have antagonistic relationship:
70. 1) Stimulus Pattern: Type of light stimulating the retina. Each column shows response for
OFF or ON cell.
71. 2) RG Firing Rate: We record retinal ganglion cells firing rate in response to light. Notice that
some cells are sustained (tonic) vs. phasic (transient).
3) Main Takeaway: The surround is always antagonist to whatever the center is doing.
4) On Center Cell: Shine light on center increase in firing with more glutamate
released/depolarization.
a. Surround Only: decrease in firing (antagonistic).
b. Highest Rate of Firing/Best Response: Shine light in center but no light in
surround.
5) Reversed for OFF cell.
a. Best Response/Maximal Firing Rate: Excited by dark spot in center and light in
surround.
6) We have parallel processing because light is seen by both an ON and OFF cell circuit and
PHASIC and TONIC receptors.
72. 1) Broad light (3) increase in response compared to baseline. This is because the response
in the center is always greater than the response in the surround.
2) Maximal firing rate in the bottom.
3) We also can really easily detect changes in light when we move from center to surround
because the firing pattern changes a LOT.
a. For example, the response of a given cell to light will be stronger if the light
portion of the visual field is adjacent to a dark portion. As a result, the retinal
mechanisms for contrast enhancement make our visual systems very sensitive to
edges or borders and allow us to perceive even weak contrasts
73. 4/14: Visual System 3
1) Off cell: Center and surround illuminated,
a. Dark middle leads to increased firing.
b. All Dark: Kind of faster than nothing because center response is stronger than
surround.
74. 1) Step Function: Moving from full light to full dark is a step function.
2) Curve Function: When we have a shadow moving across the receptive field, we are no
longer a step function.
a. B: Only cover annulus, so low rate of (antagonistic) firing.
b. C: Shadow covers middle and side leads to increased firing (center is stronger
response).
c. D: Cover all (more surround) leads to decreased firing.
3) In the curve function, we exaggerate where the edge hits, leading to an over and
undershoot.
a. Perceive huge decreases and increases as the shadow moves into the receptive field.
b. This exaggerates the edges – sharp objects (trees, shadows above you
(predator). Emphasis of contrast by the horizontal cells (center vs. surround).
75. Emphasis of Moving Objects
1) Bars of Light: On Cell
a. Reduce firing before white bar moves through the surround, increase a
lot, and decrease after it goes by.
b. Broader bar = broader peak because we illuminate center and surround
at the same time.
c. Shows exactly how wide the thing is that’s crossing our vision.
2) Parallel Processing: At the same time we have an OFF cell transmitting
complementary information better perception of image because both bright
and dark edges are accounted for.
3) Bright edges and dark edges are signaled by sharp changes in firing.
76. 4) Moving objects object elicits strong firing in the ganglion cell population near
the edges of the object’s image because these are the only regions of spatial
contrast and the only regions where the light intensity changes over time.
Color Vision:
77. 1) The L, M, and S cones are combined together gives us the spectrum of red vs. green, blue vs.
yellow.
a. Note: We don’t actually have yellow “on” blue “off” – we get it through a subtraction
process.
78. Subtypes of Retinal Ganglion Cells:
Output of Retina and Where it goes:
1) There are many types of ganglion cells mediating different types of light stimuli.
2) M-Type vs. P-Type:
a. M: Bigger cell bodies and axons, quick transmission.
i. Function: Movement in the visual field, transient responses.
b. P: Smaller axons, slower transmission, sustained response,
79. i. Function: color and texture (output of the color system). These are all in the
retina.
c. Axons of M and P type axons go to the optic nerve.
4/15: Visual Processing.
80. 1) Temporal vs. Nasal Retinal Ganglion Cells:
a. Temporal: Side portion, like the right half of the right eye.
b. Nasal: Nearer to the nose.
81.
82. 1) At the optic chiasm, the retinal ganglion cells cross to the other side of the brain.
2) Binocular Vision:
a. Differs from the frog vision. In frog vision, all the information in the right eye crosses
and goes to the left side of the brain.
b. In binocular vision, each eye is represented on both halves of the brain.
3) Blue Lines: From the right eye, the left side (nasal side) of the right retina crosses the chiasm
and goes to the opposite side of the brain.
a. From the left eye, the left side of the left eye (temporal) stays on the same side of the
brain.
b. So, we can see that both the left and right eyes send their axons to the left side of the
brain (follow blue lines).
4) Opposite for the red lines.
5) We send all these inputs into the lateral geniculate nucleus (thalamus).
83. 1) Follow the red axons. The axons from the temporal part of left eye stay on the same side of
the brain, while the nasal axons from the right eye must cross over to the left side of the
brain.
Pattern: Nasal crosses over, Temporal stays the same.
85. See I … I see, I see! (contra, ipslateral, ipslateral, contra, ips, contra)
1) The LGN has six layers, segregated by eye.
a. The axons from the ganglion cells in the retina must, in development, go to these
specific layers of the LGN.
86. b. Thus, we setup the x-y visual field by mapping this x-y visual field onto the LGN and
cortex.
2) Left Eye: The right LGN receives inputs in layers 1,4, and 6 from this contralateral left eye
through the yellow lines,
3) Right Eye: The right LGN receives inputs in layers 2,3, and 5 from the ipslateral right eye.
4) So, both eyes receive the same type of information, but inputs are segregated in the LGN by
eye.
5) Layers 1 and 2 detect motion.
a. Receive input from M-type ganglion cells (concerned with movement).
b. Land on layers 1 (contralateral) and 2 (ipslateral) viewing the same point in space.
6) Layers 3,4,5,6 very important for detecting color and texture
a. Accept input from P-type ganglion cells (smaller, for color)
LGN to Cortex (right side of diagram):
1) Layer 4 is still major input in the cortex.
2) Magnocellular channel (1,2 from LGN) serves the M cells (movement, larger) and Parvocellular
channel serves the P cells (color and texture, 3-6 from LGN).
3) All the LGN axons go to the V1 visual cortex through optic radiation (not bundled). They each go
to the cortex on their own.
4) Continual Segregation (Retinotopic Map): we saw segregation of eyes in the LGN with different
input layers. They are still separated in the cortex. For example, axons concerned with
movement (M type) versus color (P-type) land in different parts of layer IV of cortex.
a. M-Type: If we examine the two axons coding for movement (1 and 2), we see that they
represent the same point in space, but land in different places in the cortex. I.E.,
contralateral and ipslateral inputs are segregated in the cortex.
87.
88. The LGN: Stain that shows cell bodies; notice that axons go to only layers 1,4, and 6 for one
eye and 2,3, and 5 for the other eye.
89. 1) Retinotopic Map: Visual field is found on retina (x,y position), x-y mapping is maintained in LGN
(previous slide) and maintained in the visual cortex (blue and red axons land in distinct places in
the cortex).
90. A) Closer to fovea receptive field small compared to periphery (periphery has
lots of convergence).
B) Cortex dedicates a lot of space to the fovea. The fovea takes up a small area,
but is packed with cone photoceptors (1:1 ratio photoreceptor to bipolar cell)
and thus takes up almost 50% of the cortex.
C) By doing this, we get amplification of the parts of visual field that we want to
see.
91. 4/16: Final Day of Visual, Start Auditory:
V1 Cortex:
1) Layers of the V1 cortex.
2) Note that the synaptic inputs from the M and P cells go primarily to layer IV (as in the
somatosensory).
92. 1) The same point in space is seen by M cells, (Layer 4, C alpha) and P cells (Layer IV, C beta).
2) Stellate cells then transmit this information to other parts of the cortex.
3) Continual Segregation: There are still eye-specific patches in the cortex until layers 2,3. This is
where we finally get convergence of stellate cells.
Note that V1 is a lot darker/stronger because that’s where all the inputs from the LGN came in.
1) Lose x-y mapping moving from V1 and V2 parts of the cortex. Processed in the association
cortex (V2).
93. 1) Neurons project from LGN through optic radiation to the V1 cortex – they don’t travel in tight
nerves on this pathway.
2) X-Y fields from LGN are maintained and mapped retinotopically to the V1.
a. Shown in zoom in of layer four. The grey from left eye, white from the right eye in the
cortex (V1) is maintained.
See how the colors stay in specific areas of the cortex – retinotopic map idea again.
94. Input from left and right eye:
- Radioactive signal – labeled eye is where white is.
- Some particular point in x-y space is represented by the unlabeled and labelled in layer 4.
- Note the convergence after layer IV – some cells get mixed inputs (start getting binocularity)
after going past layer IV.
95. 1) In visual cortex, neurons respond selectively to bars/lines of different orientations. This comes
from alignment of many LGN center-surround receptive fields.
2) Layer IVC-Beta: Center surround responses in IVCbeta. Each cell represents a point in space.
3) Layer III (Beta): After convergence (projection neuron) in IIIbeta, we get a bar of center-
surround.
4) This “bar” of center-surround responds to specific orientations – the middle orientation.
a. There’s lots of cell nearby that responds best to different angles of light.
1) Each spot in layer 2-3 will have a cell that responds best to a specific angle of light.
96. 1) These columns consist of cells with similar functional properties.
2) Ocular Dominance: Relative strength of input from each eye.
a. Alternating Bands: Shows the segregation of left-eye and right-eye in layer IV getting
inputs from LGN.
3) Blobs: Groups of color-selective neurons – information about surfaces.
97. 4/16 and 4/18 - Auditory System:
1) The outside of the ear captures sound waves and transmits it down the ear canal (meatus) to
the sensory complex.
2) Inside the cochlea, there are axons that go to the cochlear nerve.
98. 1) As sound waves come in, the stapes drives deeper into the oval window (piston-like).
2) Stapes pushes on oval window and transduces sound through pressure changes.
a. The changes in pressure are propagated through the liquid medium (scala vestibule).
99. 1) Mechanically sensitive hair bundles protrude in the liquid inside scala media.
2) Motion in the tectorial membrane is detected by these hair cells
100. Organ of corti zoomed in.
1) Outer hair cells: Involved in amplification by changing length during basilar membrane
movement.
a. These hair cells are embedded in tectorial membrane.
2) Inner hair cells: Function is to sense frequency of sound waves.
a. Not embedded in tectorial; respond to fluid changes.
3) Afferent neurons go to the CNS.
4) Movement of tectorial and basilar membrane is detected by the hair cells.
101. 1) Characteristics of the solution in the scala media.
a. Inner hair cells are in contact with the endolymph.
b. Epithelial cells generate high K concentration inside the endolymph.
2) The high K concentrations depolarize the hair cells.
102. 1) The stereocilia attach on the top of the hair cell.
2) Hair cell is in the epithelia.
3) Cilia on top exposed to high K endolymph and the rest of cell (below) is at normal cellular
concentration of K+.
1) The cilia are structurally attached to the hair and pivot in the membrane.
2) Cell membrane actually covers the hair cells, making it possible to transduce through the hairs.
3) Takes very small amount of movement to respond to the fluid. Recall that the movement of the
two membranes (tectorial and basal) causes the fluid shift to push against the cilia. This causes
a voltage response.
103. 1) Note that there are attachments between the longer and shorter cilia.
2) Side links cause cilia to bend together.
3) After force is applied, the cilia, linked together with tip links, mechanically stretch and open
potassium channels.
104. 4) These channels transduce mechanical to electrical energy:
a. In a normal hair cell, there are about 15% of these resting potassium channels open.
b. Thus, we get potassium entering cell to cause depolarization. Resting channel is set by
this influx of ions.
1) Positive voltage outside pushes the K+ inward.
105. 1) Displacement toward tall edge opens additional channels depolarization.
2) Displacement toward short edge closes channels open at rest hyperpolarization.
3) This all occurs because of the tip link. One move, they all move together.
4) Membrane potential oscillates as we push and pull.
5) I-V relationship:
a. Lot more response when you go +200 nm vs. -200 nm.
Rectifying curve: Property that causes current to flow more readily in one direction than another. In this
case, this is caused by two things:
Two Reasons for Rectifying Curve:
Mechanical: Quickly close all the 15% of channels that were open at rest, whereas we have a decent
saturation curve on the right hand side.
Electrical: Hard to let K+ out against its concentration/voltage gradient and hyperpolarize the cell
when moving toward the short end of cilia.
b. Past 100 nm deflection all channels open, past “operating range.” Saturated.
106. Stretched cochlea.
1) Stapes at oval window transduce sound by changing pressure propagating through liquid of
scala vestibuli, basilar membrane moves in response.
107. 2) Figure C: Mechanical Operation of Basilar Membrane: basal membrane moves in response to
pressure through the oval window.
3) Figure D: Shows what really happens. Basilar membrane varies in mechanical properties. We
have differential best responses based on where we are on the membrane.
a. Apex: Thin and floppy. Responds best to low frequency.
b. Base: narrow, stiff. Responds best to high frequency.
c. Between the 20 Khz and 20 Hz, the basiliar membrane can respond best at different
positions due to properties of basiliar membrane itself.
1) Differential Best Frequency: Each wave generated reaches its max amplitude at a particular
position appropriate for the frequency of the wave, then declines in size as it goes to the apex.
a. Analogy of a wave going to shore. Max at a point, then breaks and rapidly fades.
2) Tonotopic Map: Specific arrangement of frequency-position
a. Each component of the complex sound wave establishes a wave that has its own best
frequency response at a certain position on the basilar membrane.
b. Basilar membrane is thus a mechanical frequency analyzer.
3) Example - 10K Hz: Hair cells on that part of membrane respond, membrane moves and the hair
cells depolarize rapidly too.
108. 1) Reminder: Low frequency/apex, high frequency/base.
2) Cell A: Large, slow depolarization and hyperpolarization.
a. Best Response is thus for slow frequencies (matches up).
3) Cell B: Best at high frequencies. Its depolarization and hyperpolarization is fast, so it can’t really
match with the low frequency waves too well.
a. Cell A doesn’t respond as well, though.
4) So what? This is where we start encoding stimulus frequencies and intensities!
109. 1) Transduction of Signal: Hair cells depolarize through ion channels.
a. Depolarization through K+ V-Ca channels open, Ca comes in (continual
depolarization) Ca-K channels open (K+ outward, hyperpolarize) Ca also is
pumped out reset.
110. 1) There are different transcripts of Ca-K channels in hair cells
a. Function: Responds to different frequencies.
2) Low Frequency Best Response:
a. Ca-K channel opens and closes very slowly.
3) High Frequency Best Response:
a. Different transcript (mRNA) of Ca-K channel – closes very rapidly.
4) Principle: Rate of going Ca-K open/closing governs how fast the cell can go through one cycle.
a. If we repolarize faster, we can accommodate new movement to generate a new cycle.
b. Differential distribution of Ca-K channels dictates whether cell can respond best to low
or high frequencies.
111. Antibiotics can kill hair cells! Goes through the ion channels in hair cells.
1) Outer hair cells amplify the cochlear effect.
112. 1) Auditory system is tonotopically mapped by frequency.
2) The information from one side decussates and actually goes up both sides of the CNS
3) At second synapse we’re already getting comparison of two sides of the auditory system
(the inputs)