Downloaded 403 times


































![References
• Kadmiel.M, Cidlowski.A.J, ‘Glucocorticoid Receptor Signaling in Health and Disease’ (Review).
Cell press, Trends in Pharmacological Sciences September 2013, Vol. 34, No. 9
• Nicolaides.C.N, Galata.Z, Kino.T, X, Chrousos.P.G, & Charmandari.E, ‘The Human Glucocorticoid
Receptor: Molecular Basis of Biologic Function’, PMC 2010 January 30, Volume 75(1):1,
doi:10.1016/j.steroids.2009.09.002 ( National Institute of Health, Public Access, Author
Manuscript)
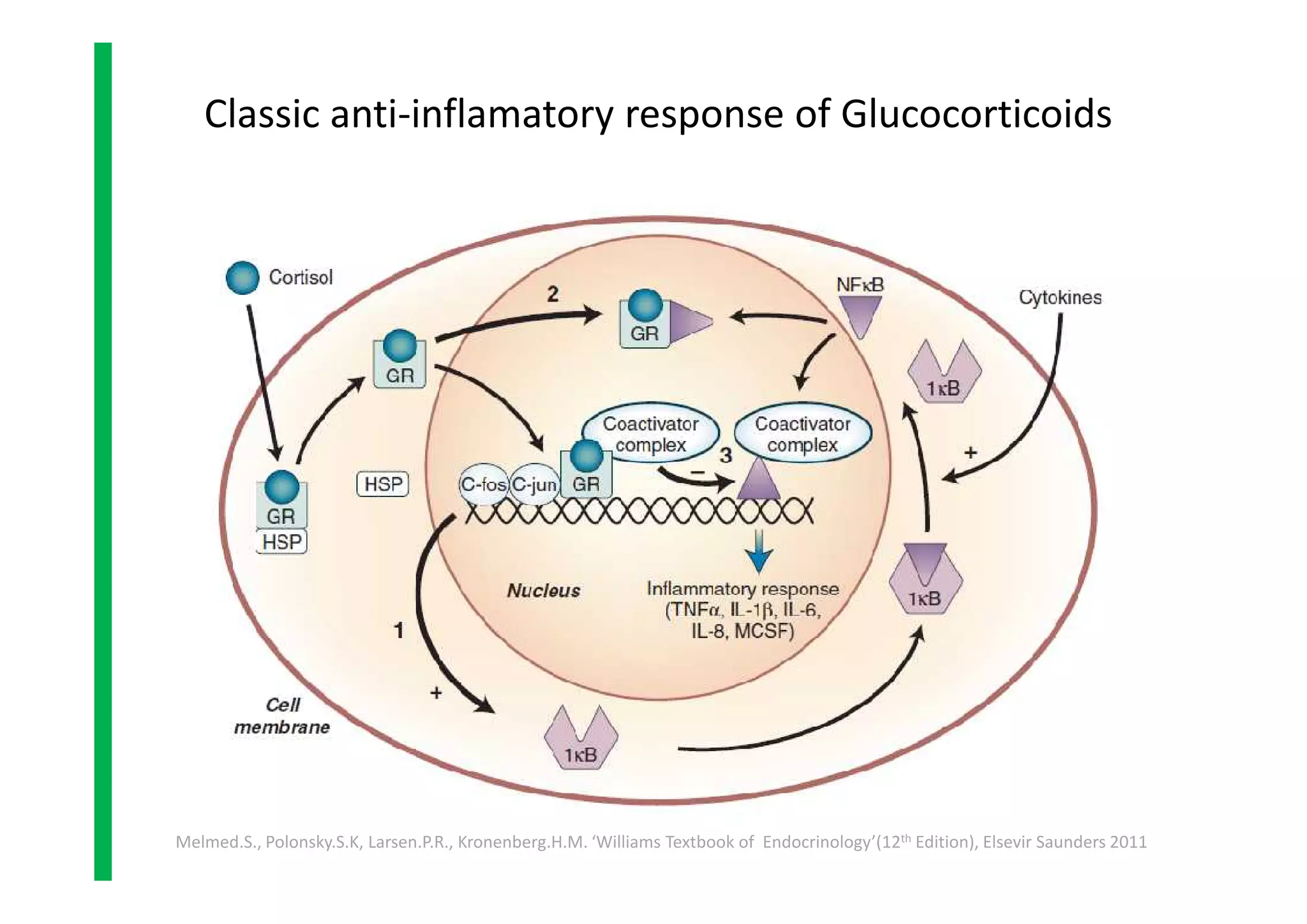
• Melmed.S., Polonsky.S.K, Larsen.P.R., Kronenberg.H.M. ‘Williams Textbook of
Endocrinology’(12th Edition), Elsevir Saunders 2011, p479-494
• Aron.C.D, Findling J.W, & Blake T.J,Edited by Garner.G.D. ‘Greenspan’s Basic & Clinical
Endocrinology,(8th Edition) ALANGE medical Book, The McGraw-Hill Companies, Inc., p346-378
• Gyton.C.A, Hall.E.J.‘Textbook of Medical Physiology’ (11th Edition), Elsevier Saunders 2006,
p944-955
• Tortora.J.Gerard, Derrickson.B, ‘Principles of Anatomy and Physiology’(13th Edition), John Wiley
& sons, Inc., p703-704
• Gottlicher M, Heck S, Herrlich P. ‘Transcriptional crosstalk, the second mode of steroid
hormone receptor action’. J Mol Med 1998;76:480–489. [PubMed: 9660166]
• Schule R, Rangarajan P, Kliewer S, Ransone LJ, Bolado J, Yang N, Verma IM, Evans RM.
‘Functional antagonism between oncoprotein c-Jun and the glucocorticoid receptor’. Cell
1990;62:1217–1226. [PubMed: 2169353]
• Jonat C, Rahmsdorf HJ, Park KK, Cato AC, Gebel S, Ponta H, Herrlich P. ‘Antitumor promotion
and anti inflammation: down-modulation of AP-1 (Fos/Jun) activity by glucocorticoid
hormone’. Cell 1990;62:1189–1204. [PubMed: 2169351]
• Kadmiel.M, Cidlowski.A.J, ‘Glucocorticoid Receptor Signaling in Health and Disease’ (Review).
Cell press, Trends in Pharmacological Sciences September 2013, Vol. 34, No. 9
• Nicolaides.C.N, Galata.Z, Kino.T, X, Chrousos.P.G, & Charmandari.E, ‘The Human Glucocorticoid
Receptor: Molecular Basis of Biologic Function’, PMC 2010 January 30, Volume 75(1):1,
doi:10.1016/j.steroids.2009.09.002 ( National Institute of Health, Public Access, Author
Manuscript)
• Melmed.S., Polonsky.S.K, Larsen.P.R., Kronenberg.H.M. ‘Williams Textbook of
Endocrinology’(12th Edition), Elsevir Saunders 2011, p479-494
• Aron.C.D, Findling J.W, & Blake T.J,Edited by Garner.G.D. ‘Greenspan’s Basic & Clinical
Endocrinology,(8th Edition) ALANGE medical Book, The McGraw-Hill Companies, Inc., p346-378
• Gyton.C.A, Hall.E.J.‘Textbook of Medical Physiology’ (11th Edition), Elsevier Saunders 2006,
p944-955
• Tortora.J.Gerard, Derrickson.B, ‘Principles of Anatomy and Physiology’(13th Edition), John Wiley
& sons, Inc., p703-704
• Gottlicher M, Heck S, Herrlich P. ‘Transcriptional crosstalk, the second mode of steroid
hormone receptor action’. J Mol Med 1998;76:480–489. [PubMed: 9660166]
• Schule R, Rangarajan P, Kliewer S, Ransone LJ, Bolado J, Yang N, Verma IM, Evans RM.
‘Functional antagonism between oncoprotein c-Jun and the glucocorticoid receptor’. Cell
1990;62:1217–1226. [PubMed: 2169353]
• Jonat C, Rahmsdorf HJ, Park KK, Cato AC, Gebel S, Ponta H, Herrlich P. ‘Antitumor promotion
and anti inflammation: down-modulation of AP-1 (Fos/Jun) activity by glucocorticoid
hormone’. Cell 1990;62:1189–1204. [PubMed: 2169351]](https://image.slidesharecdn.com/2sem-enocrinology-glucocorticoids-160302175155/75/Glucocorticoids-42-2048.jpg)


The document provides an in-depth overview of adrenal glands, detailing their anatomy, hormone production, structure of glucocorticoid receptors, and mechanisms of glucocorticoid action. Key points include the synthesis of adrenal steroid hormones, including glucocorticoids and their receptors, and the regulatory roles of various proteins in hormone action and transcriptional activation. Furthermore, it outlines the significance of the adrenal cortex and the physiological impacts of glucocorticoids in the body.











































































































