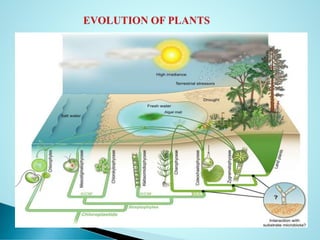
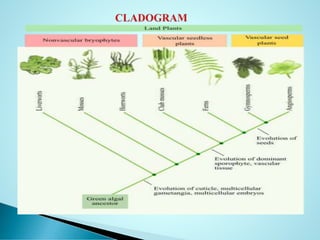
Angiosperms or flowering plants, comprising about 300,000 species, emerged in the Cretaceous period, dominating vegetative life and adapting to diverse habitats. The document discusses their evolutionary characteristics, including the structure of flowers, the process of double fertilization, and the debate over their origin, with theories suggesting either a monophyletic or polyphyletic ancestry. Key differences between homology and analogy in evolution are also explored, alongside the development of phylogenetic trees that illustrate relationships among plant groups.