3
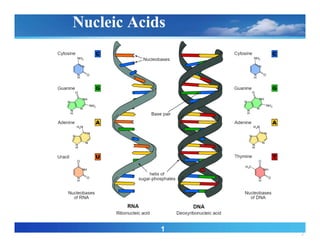
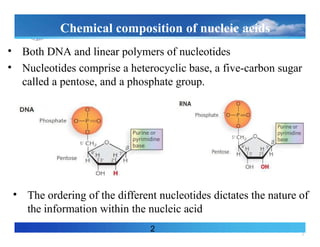
Chemical composition ofnucleic acids
• Both DNA and linear polymers of nucleotides
• Nucleotides comprise a heterocyclic base, a five-carbon sugar
called a pentose, and a phosphate group.
2
• The ordering of the different nucleotides dictates the nature of
the information within the nucleic acid
4.
4
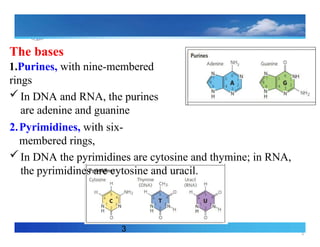
The bases
3
1.Purines, withnine-membered
rings
In DNA and RNA, the purines
are adenine and guanine
2.Pyrimidines, with six-
membered rings,
In DNA the pyrimidines are cytosine and thymine; in RNA,
the pyrimidines are cytosine and uracil.
5.
5
The sugars ofnucleic acids
• In DNA, the sugar is always deoxyribose; whereas, in RNA, it
is ribose.
• Deoxyribose has one less oxygen than ribose. It is this
chemical difference that gave rise to the names
deoxyribonucleic acid and ribonucleic acid.
• The oxygen atom in 2’ position in RNA nucleotide makes it more reactive
and less chemically stable than DNA. Thus, DNA is better suited to serve
as the long term
‐ repository of genetic information.
6.
6
5
The Phosphate group
•consists of a phosphorus atom bonded to four oxygen atoms
• carrying a negative charge, which makes DNA acidic hence
nucleic acid.
• always bonded to the 5`‐ carbon atom of the sugar in a
nucleotide
The negatively charged phosphates are extremely insoluble in
lipids. Thus, the insolubility of nucleic acids in lipids ensures that
they are retained within the cell
7.
7
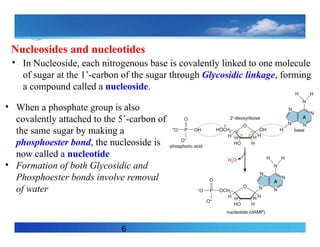
Nucleosides and nucleotides
6
•In Nucleoside, each nitrogenous base is covalently linked to one molecule
of sugar at the 1’-carbon of the sugar through Glycosidic linkage, forming
a compound called a nucleoside.
• When a phosphate group is also
covalently attached to the 5’-carbon of
the same sugar by making a
phosphoester bond, the nucleoside is
now called a nucleotide
• Formation of both Glycosidic and
Phosphoester bonds involve removal
of water
8.
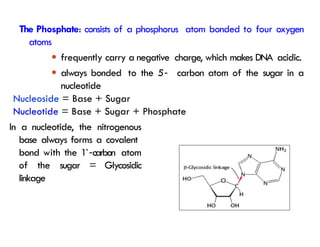
In a nucleotide,the nitrogenous
base always forms a covalent
bond with the 1`‐carbon atom
of the sugar = Glycosidic
linkage
The Phosphate: consists of a phosphorus atom bonded to four oxygen
atoms
● frequently carry a negative charge, which makes DNA acidic.
● always bonded to the 5
`‐ carbon atom of the sugar in a
nucleotide
Nucleoside = Base + Sugar
Nucleotide = Base + Sugar + Phosphate
9.
9
Nucleotide Function
• Buildingblocks for DNA and RNA
• Intracellular source of energy - Adenosine triphosphate (ATP)
• Enzyme cofactors (NAD+)
• Signal transduction: Second messengers - Involved in
intracellular signaling (e.g. cyclic adenosine monophosphate
[cAMP])
10.
10
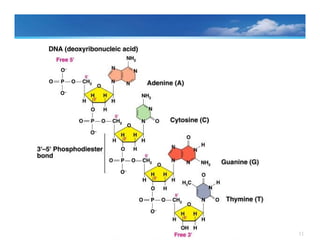
Structure of theDNA
• Polynucleotide chain are formed when the 3’-hydroxyl of one
nucleotide and the 5’-phosphate are attached through a
phosphodiester linkage
• Phosphodiester linkages
create the repeating, sugar– phosphate backbone of the
polynucleotide chain
impart an inherent polarity to the DNA chain (a free 5’-
phosphate at one end and a free 3’-hydroxyl at the other end)
7
12
Significance of Nucleicacid polarity (5’ and
3’)
• Understanding NA polarity is critical for
understanding aspects of replication and transcription
reading a DNA sequence, and carrying out experiments in the
lab.
• By convention, a NA sequence is written with the 5’ end to the
left and the 3’end to the right. This makes sense because 5’→ 3’is
the direction of synthesis for both DNA and RNA.
Example: 5`ATAAGCTC3`, or even just ATAAGCTC.
• Note that the directionality of the chain means that, for example,
ATAAG is not the same asGAATA
8
13.
The structure ofDNA: the double helix
`
● DNA most commonly occurs in nature as the well‐known
‘double helix’.
● deduced by James Watson and Francis Crick in 1953.
• DNA structure is a right handed double
‐
helix.
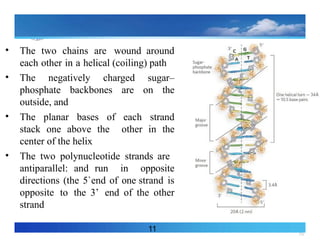
• The two chains are wound around each
other a helical (coiling) path
• The negatively charged sugar–
phosphate backbones are on the
outside, and
• The planar bases of each strand stack
one above the other in the center of
the helix
14.
14
DNA Molecules HaveDistinctive Base Compositions
• In the 1940s, Erwin Chargaff and his colleagues made an
important discovery that provided clues to the structure of DNA.
1. The base composition of DNA generally varies from one species
to another
2. DNA specimens isolated from different tissues of the same
species have the same base composition.
3. The base composition of DNA in a given species does not change
with an organism’s age, nutritional state, or environment.
4. In all cellular DNAs, regardless of the species, the number of
A=T and the number of G= C. From these A+G = T + C.
Chargaff’s rules
t 9
The structure of DNA: the
doublehelix
15.
15
The structure ofDNA: the
doublehelix
`
• DNA most commonly occurs in nature as the well known
‐
‘double helix’.
• deduced by James Watson and Francis Crick in 1953.
• DNA structure is a right handed
‐ double helix.
16.
16
• The twochains are wound around
each other in a helical (coiling) path
• The negatively charged sugar–
phosphate backbones are on the
outside, and
• The planar bases of each strand
stack one above the other in the
center of the helix
• The two polynucleotide strands are
antiparallel: and run in opposite
directions (the 5`end of one strand is
opposite to the 3’ end of the other
strand
11
17.
17
12
• there aretwo grooves that
run
the
the backbone of
the major and
minor
between
strands:
grooves
The major groove has a
significant role in sequence-
specific DNA-protein
interactions.
Proteins that carry out their function
by binding DNA and read the string
of base letters exposed in the groove
18.
18
13
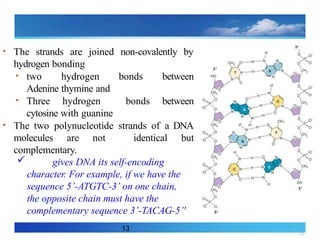
• The strandsare joined non-covalently by
hydrogen bonding
• two hydrogen bonds between
Adenine thymine and
• Three hydrogen bonds between
cytosine with guanine
• The two polynucleotide strands of a DNA
molecules are not identical but
complementary.
gives DNA its self-encoding
character. For example, if we have the
sequence 5’-ATGTC-3’ on one chain,
the opposite chain must have the
complementary sequence 3’-TACAG-5’'
20
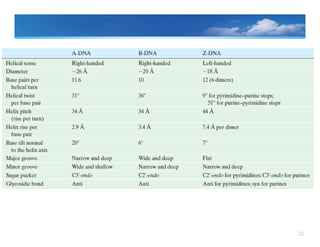
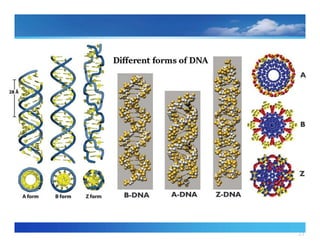
Variations in thestructureof DNA (A, B, Z)
The precise three dimensional
‐ shape of the DNA molecule can vary,
depending on the conditions in which the DNAis placed and on the base
sequence itself.
B Form
• Described by Watson and Crick
• exists when plenty of water surrounds the
DNA molecule ( at high Humidity)
• Has no unusual base sequence in the DNA
• is the moststable configuration
and predominant structure in the cell
• possesses approximately 10.5 base pairs
(bp) per turn of the helix;
21.
21
A‐ Form
• existswhen le s water is present.
• Is also alpha helical (right handed)
‐ but shorter and wider than
Bform
• There is little evidence that A form exist
under conditions.
Z form
‐
• forms a left handed helix
physiological
• Its sugar–phosphate backbones zigzag back and forth, giving rise to
the name Z form (for zigzag)
• can arise under physiological conditions when particular base
sequences are present, such as stretches of alternating C and G
sequences
24
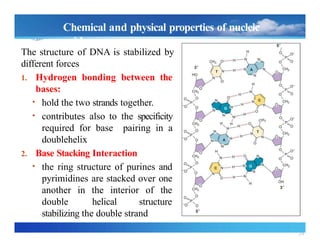
Chemical and physicalproperties of nucleic
acids
The structure of DNA is stabilized by
different forces
1. Hydrogen bonding between the
bases:
• hold the two strands together.
• contributes also to the specificity
required for base pairing in a
doublehelix
2. Base Stacking Interaction
• the ring structure of purines and
pyrimidines are stacked over one
another in the interior of the
double helical structure
stabilizing the double strand
25.
25
Effects of acidsand bases on the structure of DNA
• Strong acid and high temperatures hydrolyzed the DNA completely
to their constituents
• Dilute mineral acid (for example pH 3–4) break down the
glycosylic bonds attaching the purine bases to the ribose ring, and
hence the nucleic acid becomes apurinic
• Increasing pH above the physiological range break down the
double stranded structure of the DNA(denatured)
Thermal properties of DNA
1. UV absorption
• Nucleic acids absorb UV light due to the conjugated aromatic
nature of the bases at 260nm (λmax=260nm), which is conveniently
distinct from the λmax of protein (280nm)
• can be used for detection, quantitation and a se sment of DNApurity
26.
26
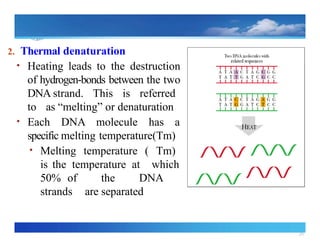
2. Thermal denaturation
•Heating leads to the destruction
of hydrogen bonds
‐ between the two
DNA strand. This is referred
to as “melting” or denaturation
• Each DNA molecule has a
specific melting temperature(Tm)
• Melting temperature ( Tm)
is the temperature at which
50% of the DNA
strands are separated
27.
27
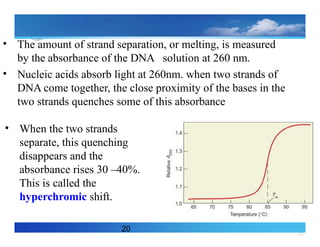
20
• The amountof strand separation, or melting, is measured
by the absorbance of the DNA solution at 260 nm.
• Nucleic acids absorb light at 260nm. when two strands of
DNA come together, the close proximity of the bases in the
two strands quenches some of this absorbance
• When the two strands
separate, this quenching
disappears and the
absorbance rises 30 –40%.
This is called the
hyperchromic shift.
28.
28
The melting temperatureis affected by different factors
i. The pH and salt concentration of the solution.
• Extremes of pH disrupt hydrogen bonds .
• Higher ionic concentrations suppress the electrostatic repulsion
between the negatively charged phosphate groups on the
backbone and hence exert a stabilizing effect.
• In pure water, DNA can melt even at room temperature
i. Base Composition
• The higher the proportion of GCbase pairs, the higher the
melting temperature. ATbase pairs are weaker, as they have only
two hydrogen bonds, opposed to GCpairs, which havethree
hydrogen bonds
• DNA base compositions are often cited as the GCratio.
• The GCcontent (% G +C) is calculated from the fractional composition
of bases as follows:
29.
29
Renaturation
• The thermaldenaturation of DNA may be
reversed by cooling the solution.
• The renaturation of the regions of
complementary nucleic acid strands is
known as hybridization.
• The rate of renaturation depends on the
temperature, cooling time and Concentration
of the DNA solution
The best temperature for
renaturationof a DNA is about 250C
below its Tm
the higher the concentration, the faster the
rate of renaturation
the longer the time a lowed for annealing,
the more wi l it occur
30.
30
DNA
Supercoiling
• Double strandedDNA may be either linear or circular.
• Circular DNAhas no ends.
Mitochondrial DNA and the DNA of most prokaryotes
and some viruses are closed circular structures
• Circular DNA may exist in relaxed form (no supercoils) or
it may be converted to more compact structures by super
coiling
• Supercoiling means the coiling of a coil.
• DNA is coiled in the form of a double helix, with both
strands of the DNA coiling around an axis. The further
coiling of that axis upon itself produces DNAsupercoiling.
31.
31
www.hu.edu.et Ever toExcel!
27
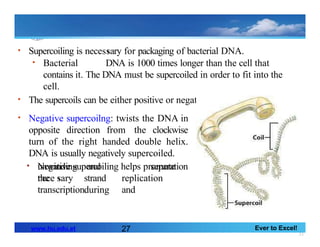
• Supercoiling is necessary for packaging of bacterial DNA.
• Bacterial DNA is 1000 times longer than the cell that
contains it. The DNA must be supercoiled in order to fit into the
cell.
• The supercoils can be either positive or negative
• Negative supercoilng: twists the DNA in
opposite direction from the clockwise
turn of the right handed double helix.
DNA is usually negatively supercoiled.
• Negative supercoiling helps promote
the
separation
unwinding
nece sary
and
strand
during
replication
and
transcription.
32.
32
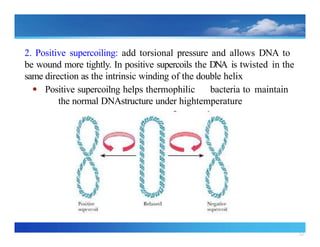
2. Positive supercoiling:add torsional pressure and allows DNA to
be wound more tightly. In positive supercoils the DNA is twisted in the
same direction as the intrinsic winding of the double helix
● Positive supercoilng helps thermophilic bacteria to maintain
the normal DNAstructure under hightemperature
33.
33
Topoisomerases and DNAGyrase
•The total amount of twisting present in a
DNA molecule is referred to as the
linking number (L). This is the sum
of the contributions due to the double
helix plus the super coiling.
‐
• The number of double helical turns is
sometimes known as the twist, T, and
the number of supercoils in DNA as the
writhe or writhing number, W
. In this
terminology, the linking number, L, is
the sum of the twist plus the
writhe(L=T+W).]
34.
• The samecircular DNA molecule can have different numbers of
supercoils. These forms are known as topological isomers, or
topoisomers.
• The enzymes that insert or remove supercoils are
therefore named topoisomerases.
Type I topoisomerases
• break only one strand of DNA, which changes the linking number in
steps of one.
Type II topoisomerases
• break both strands of the DNA and pass another part of the double helix
through the gap. This changes the linking number in steps of two
• DNA gyrase, is a type II topoisomerase, that introduce negative
supercoils
• into closed circular molecules of DNA, such as plasmids or the bacterial
chromosome.
34
35.
35
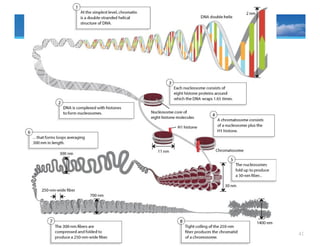
• Plants andanimals have vastly more DNA than bacteria and must fold this
DNA to fit into the cell nucleus.
• Eukaryotic chromosomes may be as much as a centimeter long and must be
folded up to fit into the cell nucleus, which is five microns across, a
necessity for a 2,000 fold shortening.
• The mechanism of packaging involves winding the DNA around special
proteins called histones
• Eukaryotic cells contain 5 kinds of histones, H1, H2A, H2B, H3 and H4
• Histones have a high percentage of arginine and lysine
• give them a net positive charge.
• The positive charges attract the negative charges on the phosphates of DNA
and holds the DNA in contact with the histones.
DNA packaging in Eukaryotes
36.
36
H1- a linkerhistone binds to the
nucleosome and stabilize the point at
which the DNA enters and leaves
the nucleosome core.
form the 10 nm fiber
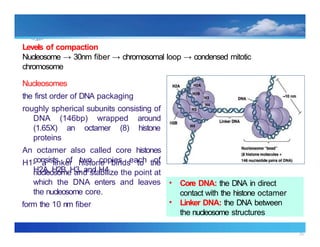
Levels of compaction
Nucleosome → 30nm fiber → chromosomal loop → condensed mitotic
chromosome
Nucleosomes
the first order of DNA packaging
roughly spherical subunits consisting of
DNA (146bp) wrapped around
(1.65X) an octamer (8) histone
proteins
An octamer also called core histones
consists of two copies each of
H2A, H2B, H3, and H4
• Core DNA: the DNA in direct
contact with the histone octamer
• Linker DNA: the DNA between
the nucleosome structures
38
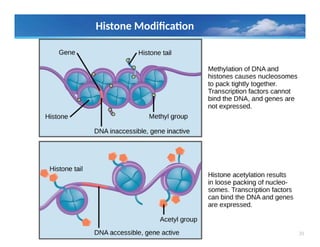
Histone Modification
• Allof the histones are subject to numerous covalent
modifications, most of which occur in the histone tails.
• The histone tails can be acetylated, methylated, phosphorylated,
and ubiquitylated at numerous sites
• All of these modifications are reversible, and a given
modification might exist only transiently, or can be maintained
stably through multiple cell divisions.
• Modification of histones is associated with structural changes that
occur in chromatin at replication and transcription, and
specific modifications also facilitate DNA repair.
• Modifications at specific positions on specific histones can define
different functional states of chromatin.
40
• The nucleosomeprovides the first level of organization, compacting the
DNA about 6-fold over the length of naked DNA, resulting in a “beads-on-
a-string” fiber of approximately 10 nm in diameter.
• The secondary level of organization involves interactions between
nucleosomes of the 10-nm fiber, leading to more condensed chromatin
fibers.
This fiber is of approximately 30 nm in diameter. The structure of this
fiber requires the histone tails and is stabilized by linker histones.
• The final, tertiary level of chromatin organization requires the further
folding and compacting of chromatin fibers into the 3D structures of
interphase chromatin or mitotic chromosomes.
42
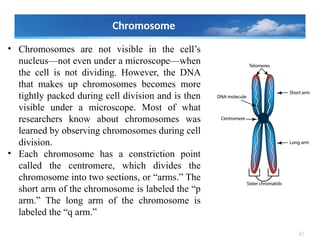
• Chromosomes arenot visible in the cell’s
nucleus—not even under a microscope—when
the cell is not dividing. However, the DNA
that makes up chromosomes becomes more
tightly packed during cell division and is then
visible under a microscope. Most of what
researchers know about chromosomes was
learned by observing chromosomes during cell
division.
• Each chromosome has a constriction point
called the centromere, which divides the
chromosome into two sections, or “arms.” The
short arm of the chromosome is labeled the “p
arm.” The long arm of the chromosome is
labeled the “q arm.”
Chromosome
43.
43
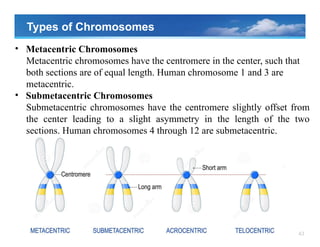
Types of Chromosomes
•Metacentric Chromosomes
Metacentric chromosomes have the centromere in the center, such that
both sections are of equal length. Human chromosome 1 and 3 are
metacentric.
• Submetacentric Chromosomes
Submetacentric chromosomes have the centromere slightly offset from
the center leading to a slight asymmetry in the length of the two
sections. Human chromosomes 4 through 12 are submetacentric.
44.
44
• Acrocentric Chromosomes
Acrocentricchromosomes have a centromere which is severely offset
from the center leading to one very long and one very short section.
Human chromosomes 13,15, 21, and 22 are acrocentric.
• Telocentric Chromosomes
Telocentric chromosomes have the centromere at the very end of the
chromosome. Humans do not possess telocentric chromosomes but they
are found in other species such as mice.
Types of Chromosomes...




![9
Nucleotide Function
• Building blocks for DNA and RNA
• Intracellular source of energy - Adenosine triphosphate (ATP)
• Enzyme cofactors (NAD+)
• Signal transduction: Second messengers - Involved in
intracellular signaling (e.g. cyclic adenosine monophosphate
[cAMP])](https://image.slidesharecdn.com/molbiollecture2b-250220160216-8cefc5e7/85/Mol_Biol_Lecture-2b-pptxjgctddgdsefdfxdxe-9-320.jpg)













![33
Topoisomerases and DNAGyrase
• The total amount of twisting present in a
DNA molecule is referred to as the
linking number (L). This is the sum
of the contributions due to the double
helix plus the super coiling.
‐
• The number of double helical turns is
sometimes known as the twist, T, and
the number of supercoils in DNA as the
writhe or writhing number, W
. In this
terminology, the linking number, L, is
the sum of the twist plus the
writhe(L=T+W).]](https://image.slidesharecdn.com/molbiollecture2b-250220160216-8cefc5e7/85/Mol_Biol_Lecture-2b-pptxjgctddgdsefdfxdxe-33-320.jpg)


































































![Polymer [ बहुलक ] Chemistry Notes PDF - Irfanullah Mehar - JJ Sir Chemistry.pdf](https://cdn.slidesharecdn.com/ss_thumbnails/polymerchemistrynotespdf-irfanullahmehar-jjsirchemistry-260210172118-3f9b37f7-thumbnail.jpg?width=640&height=640&fit=bounds)






