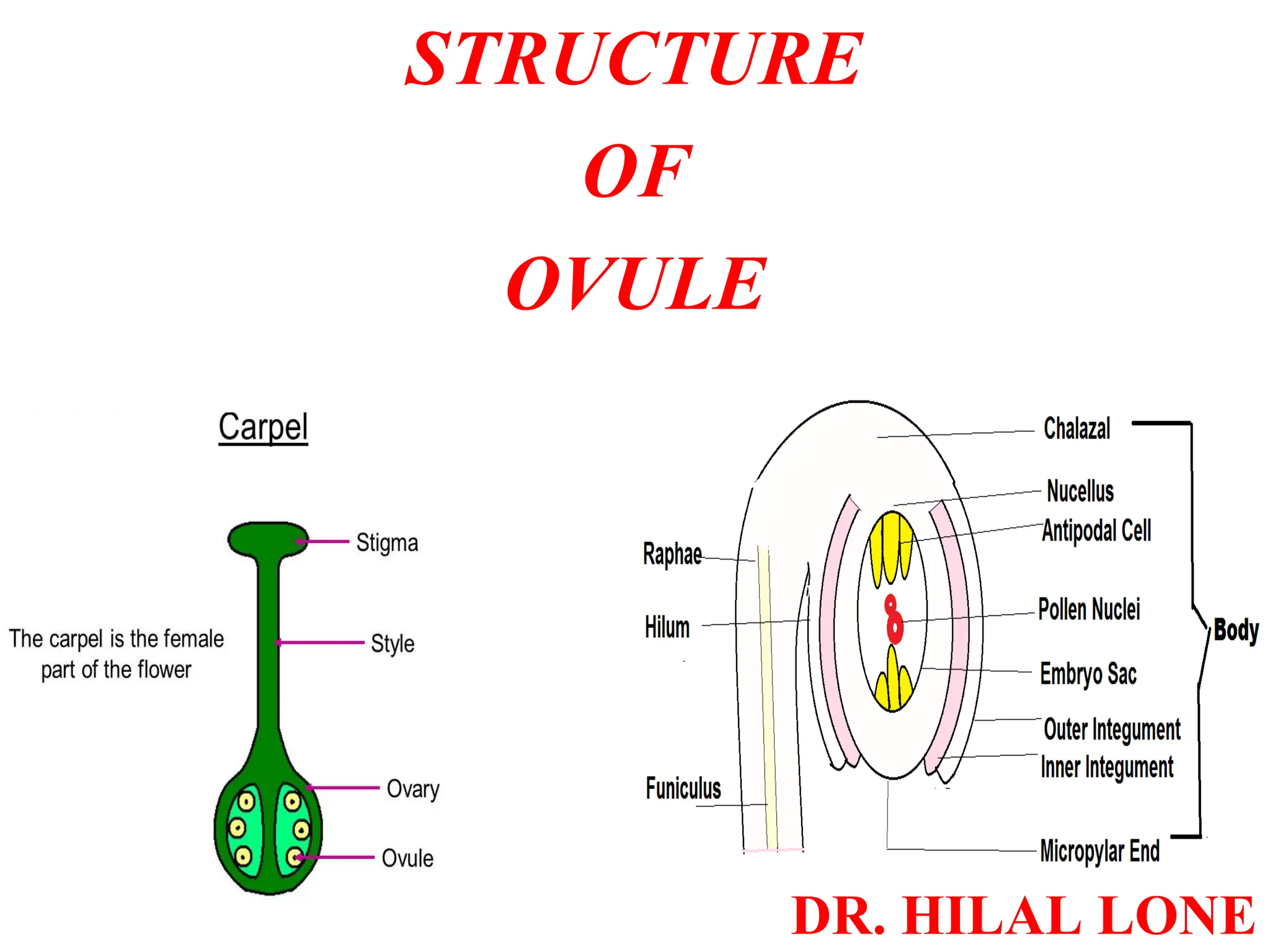
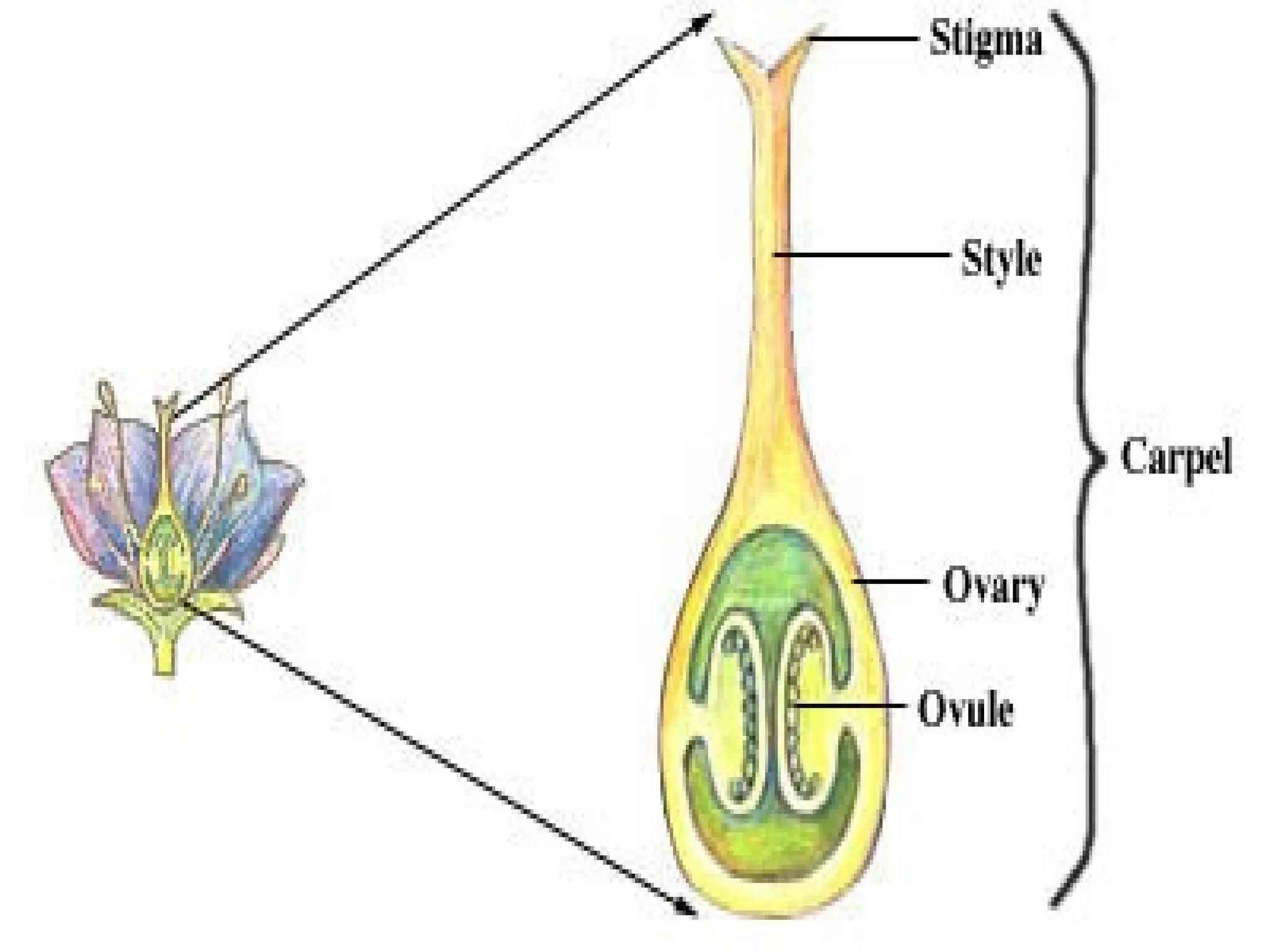
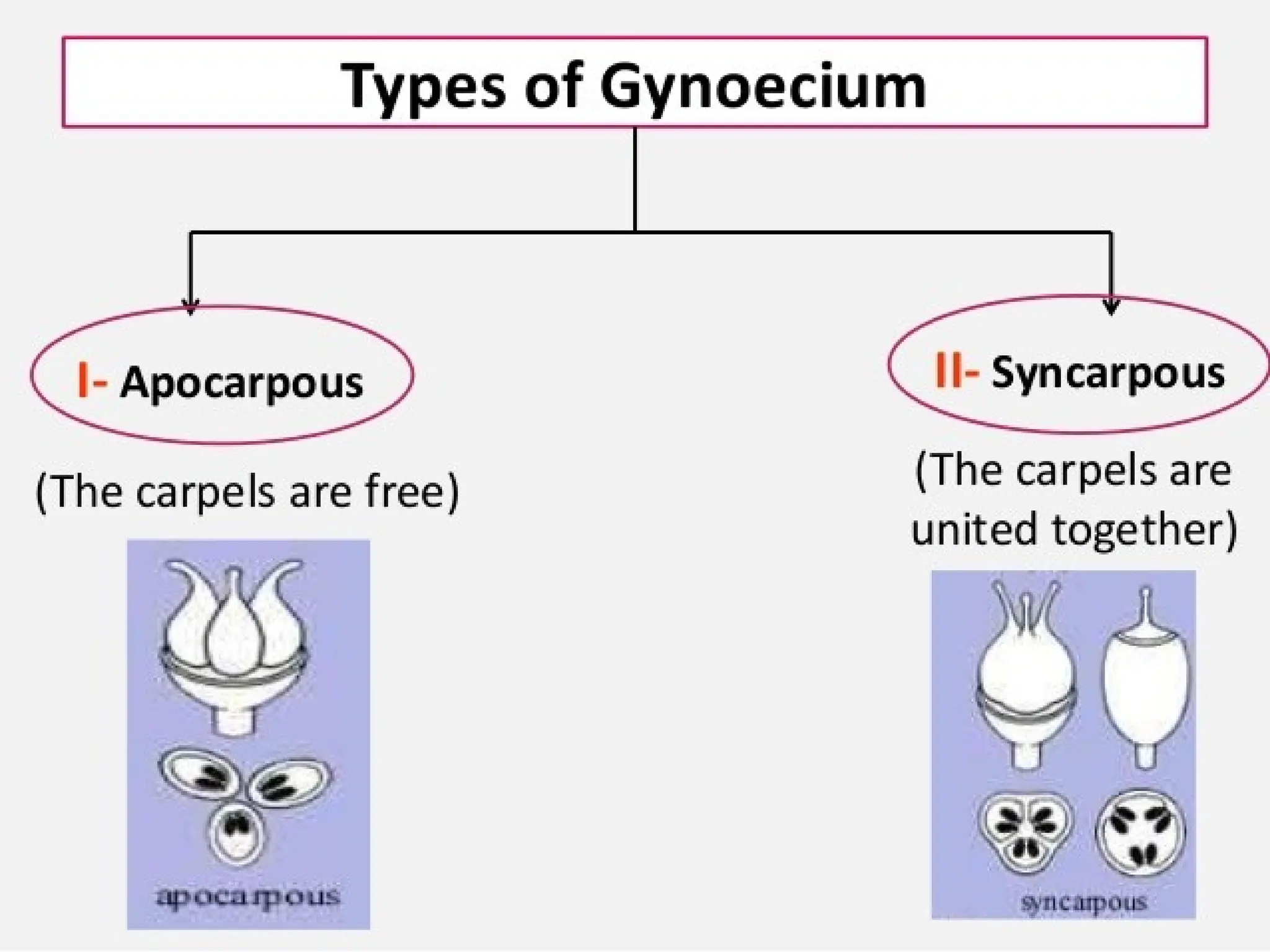
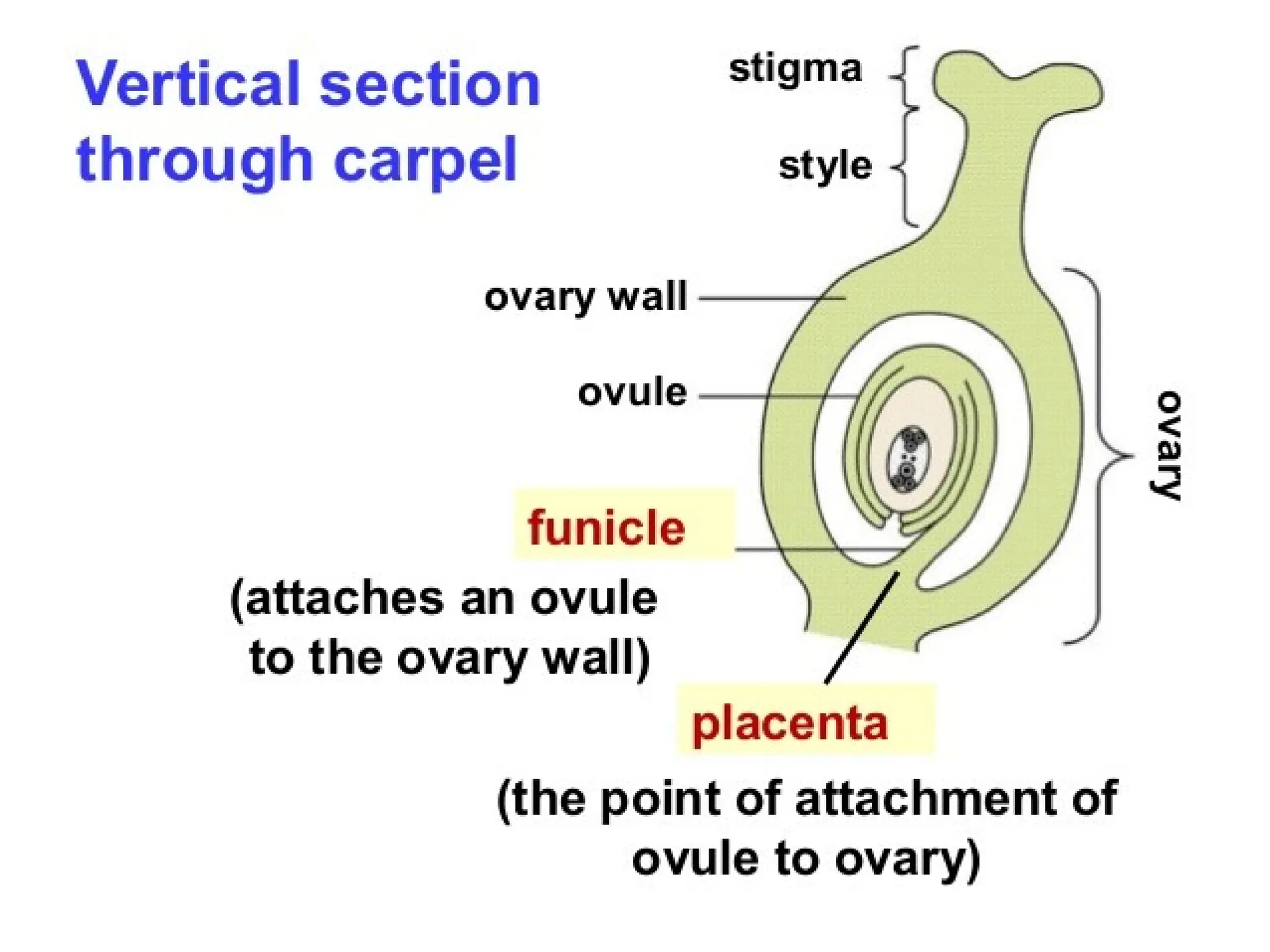
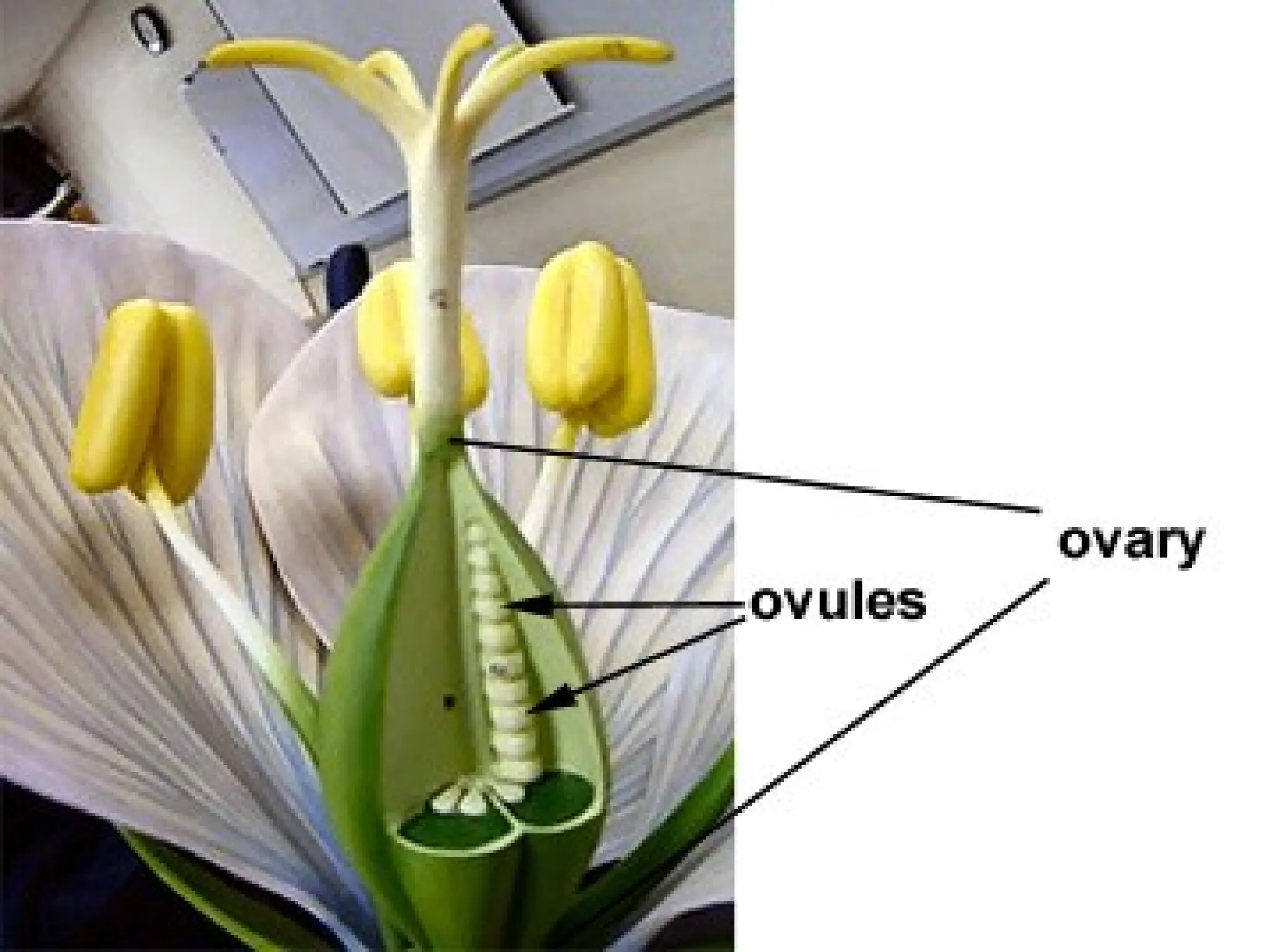
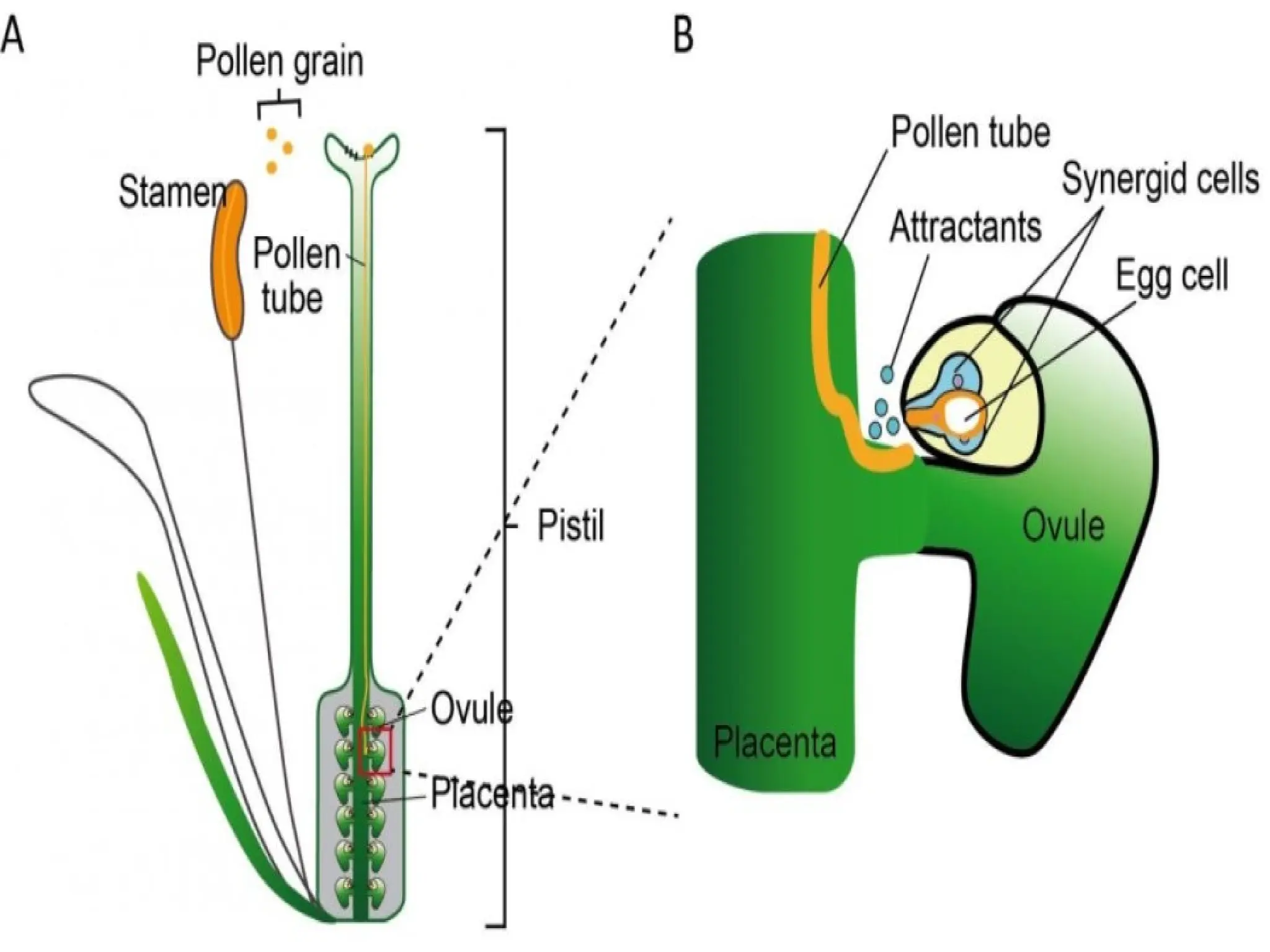
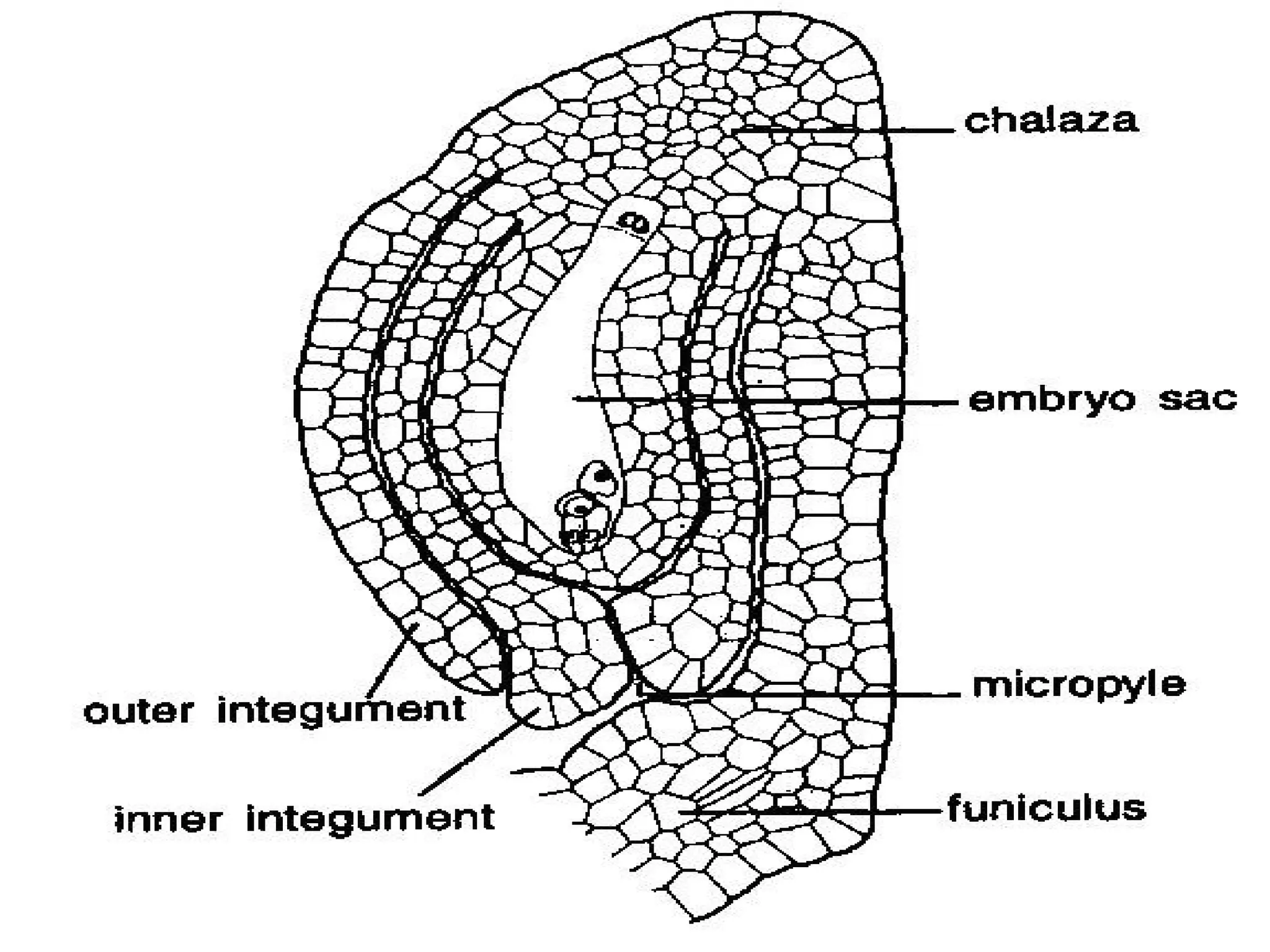
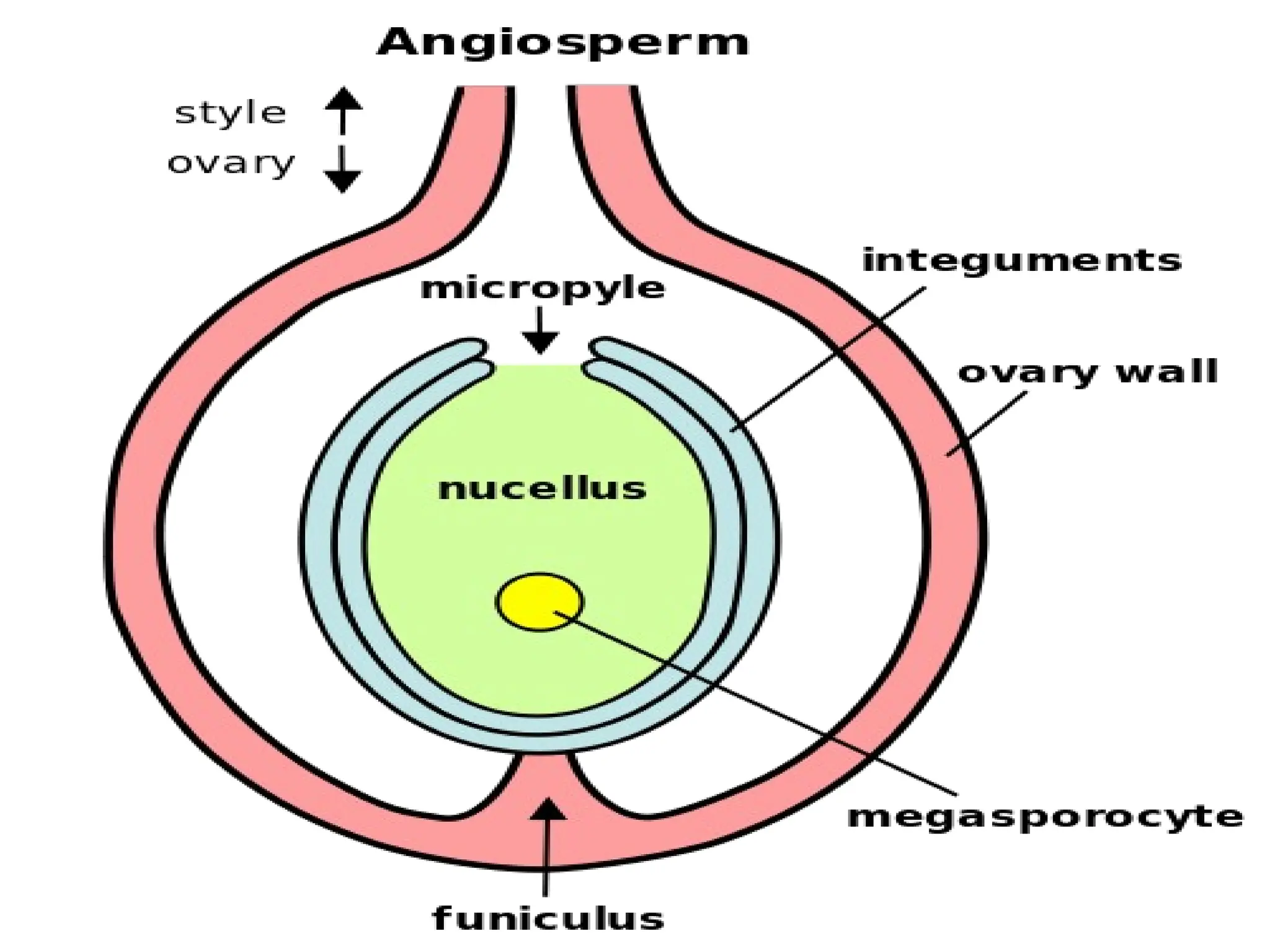
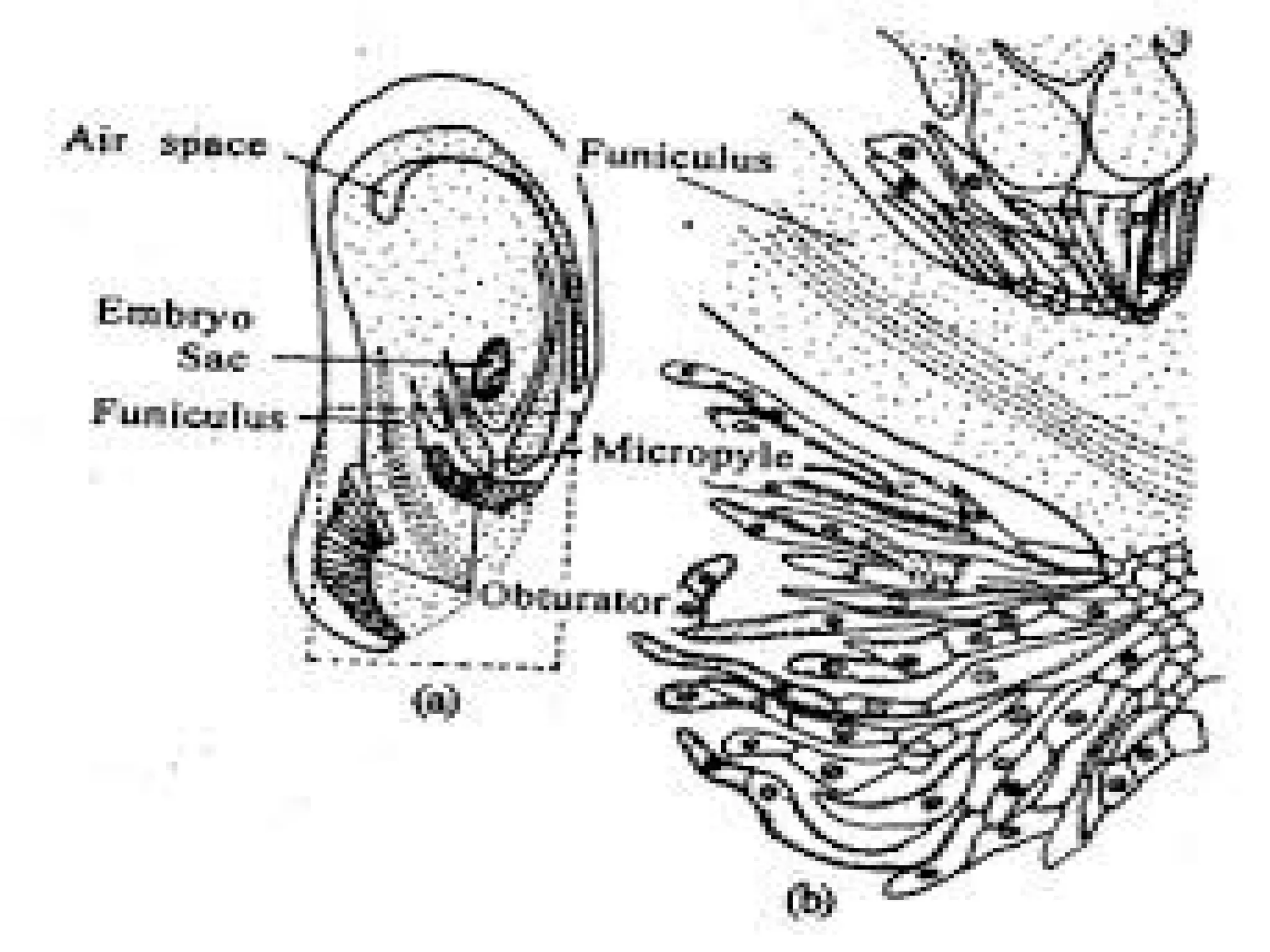
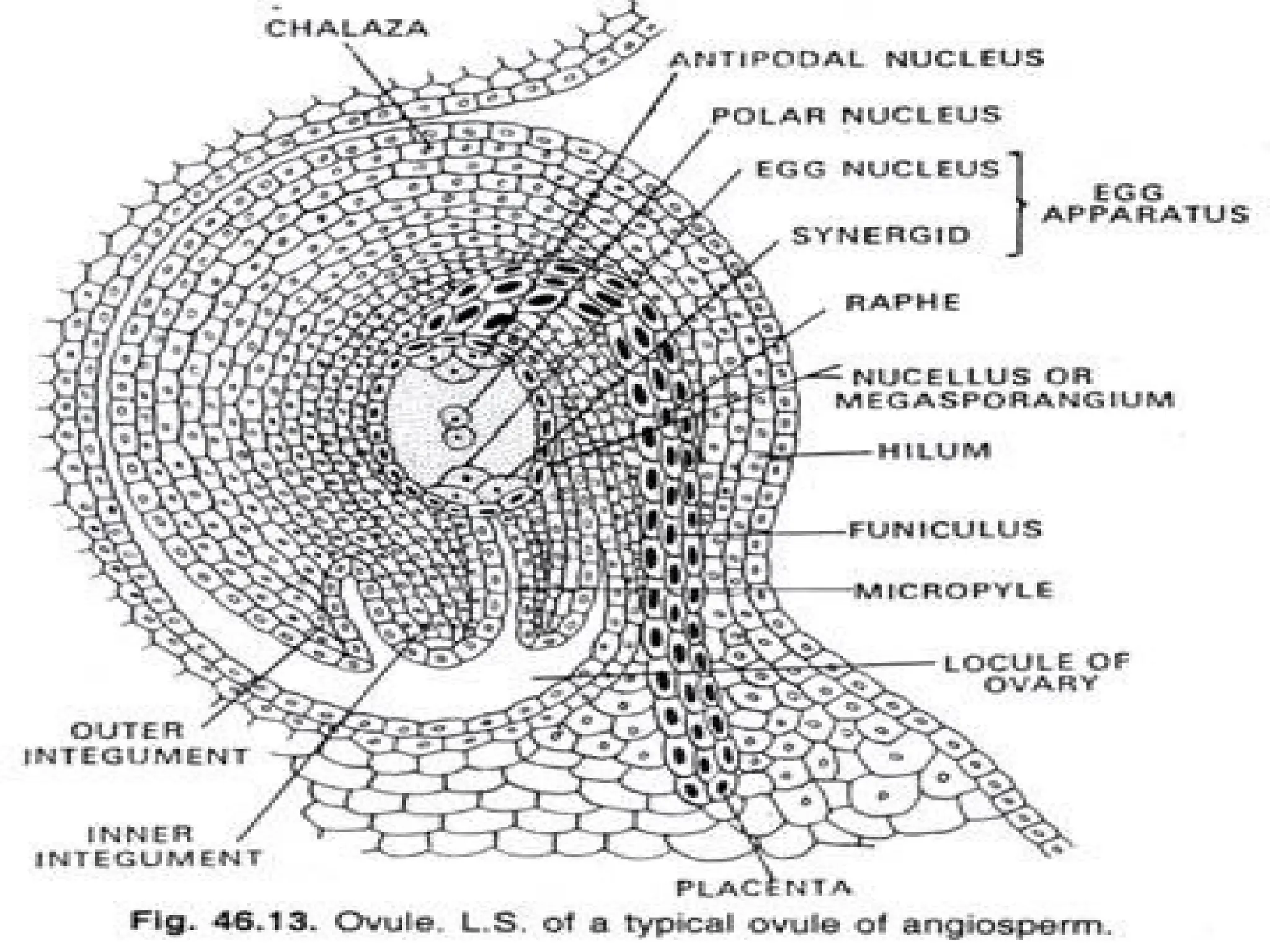
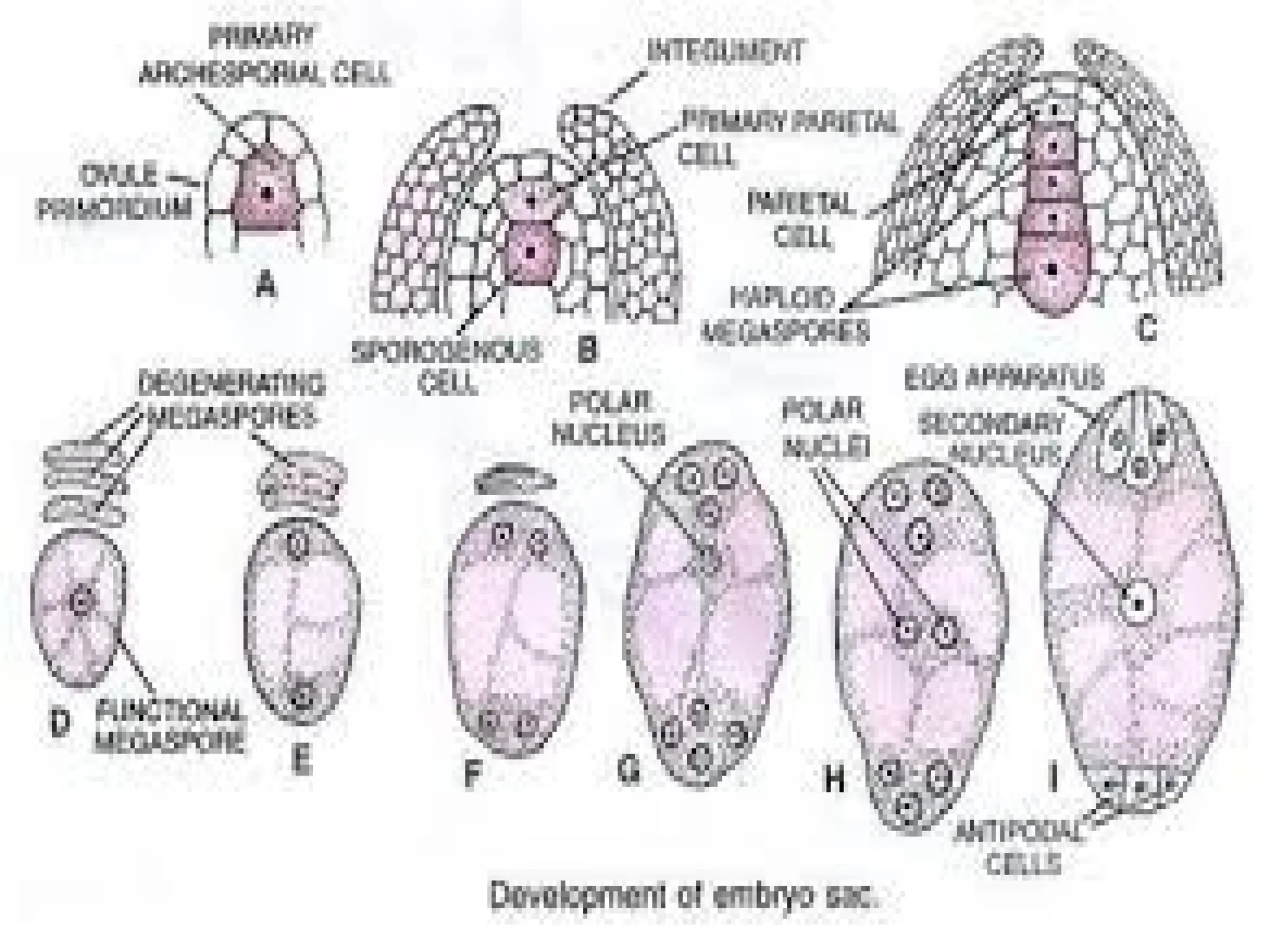
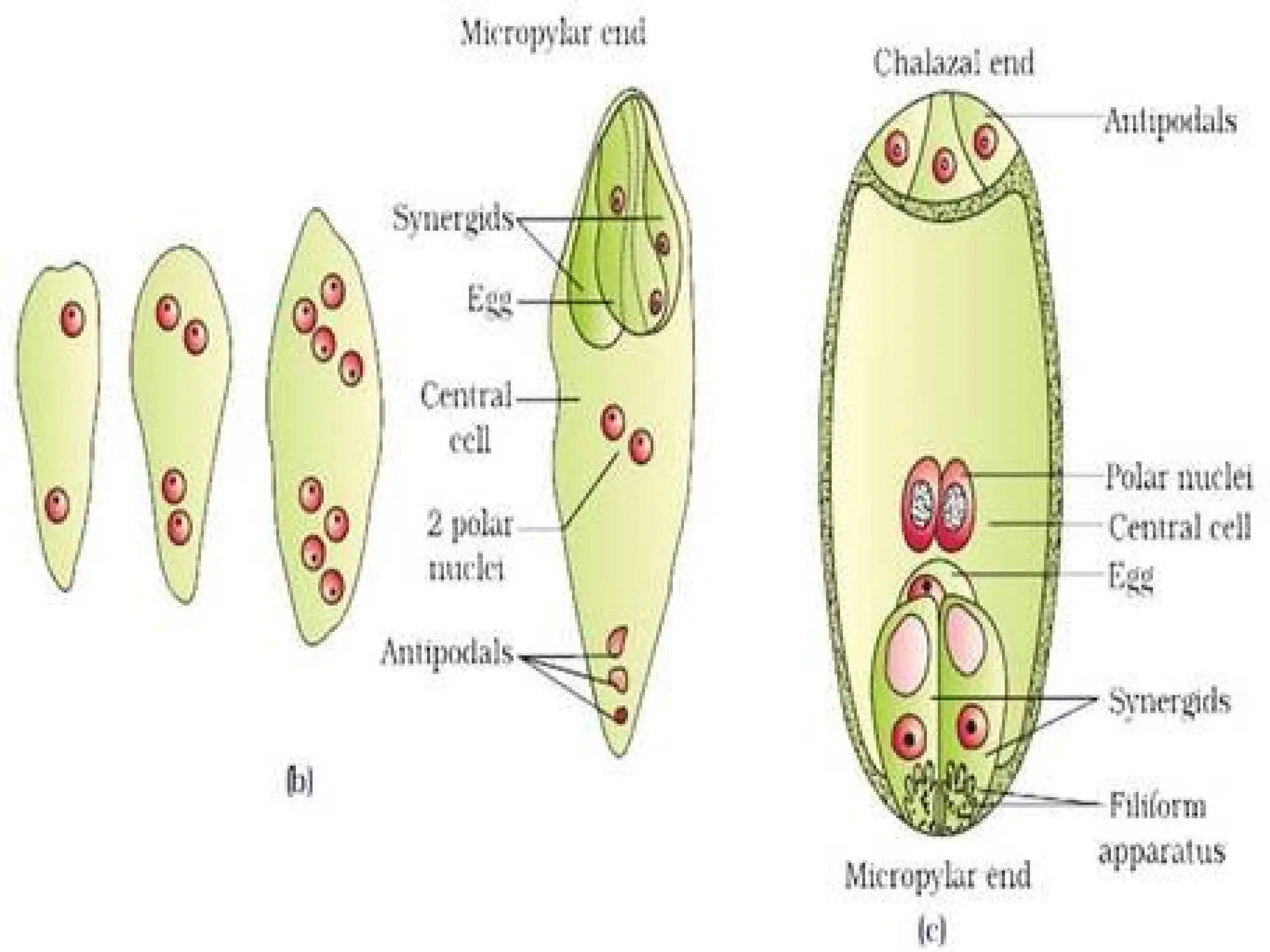
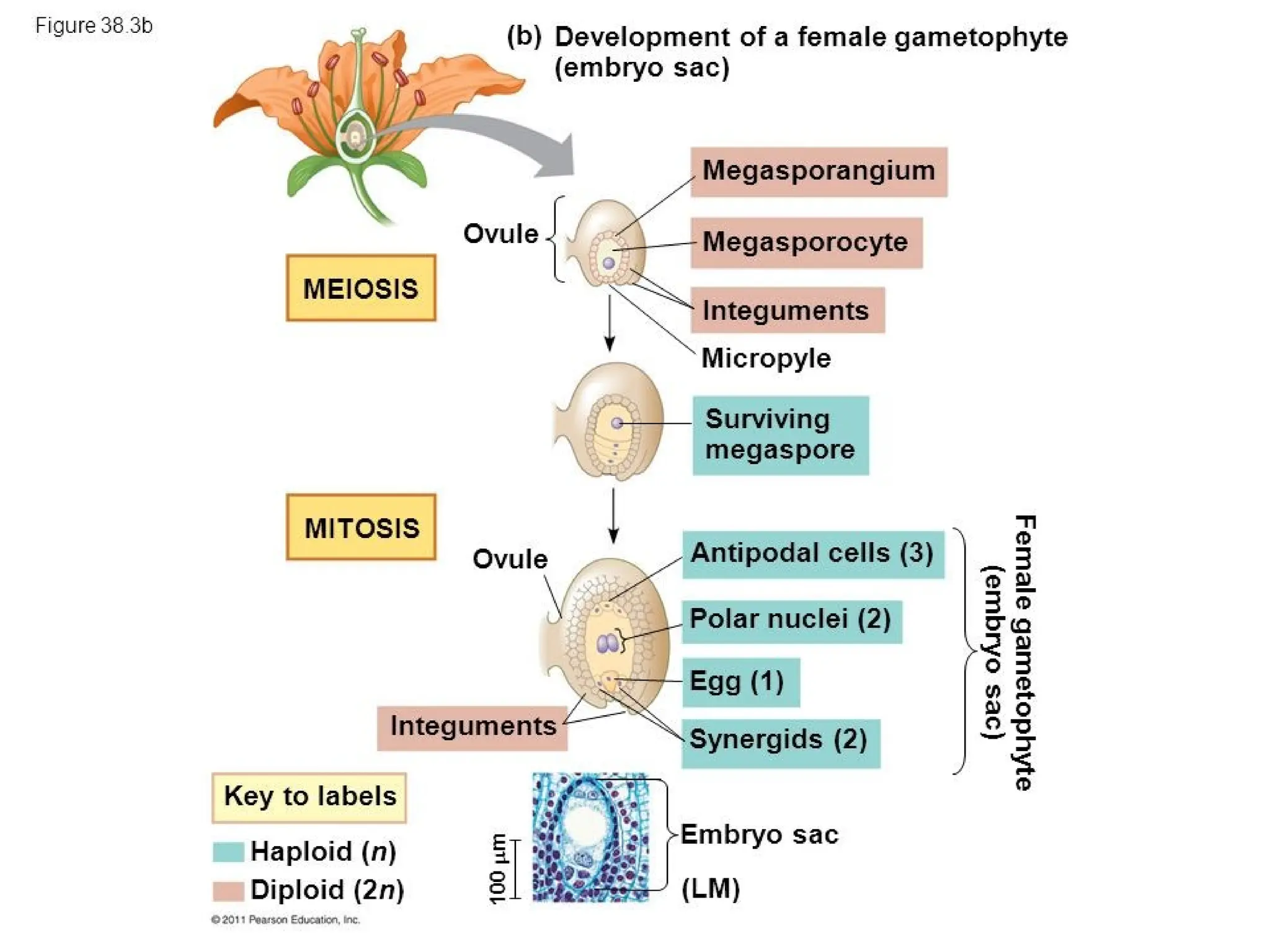
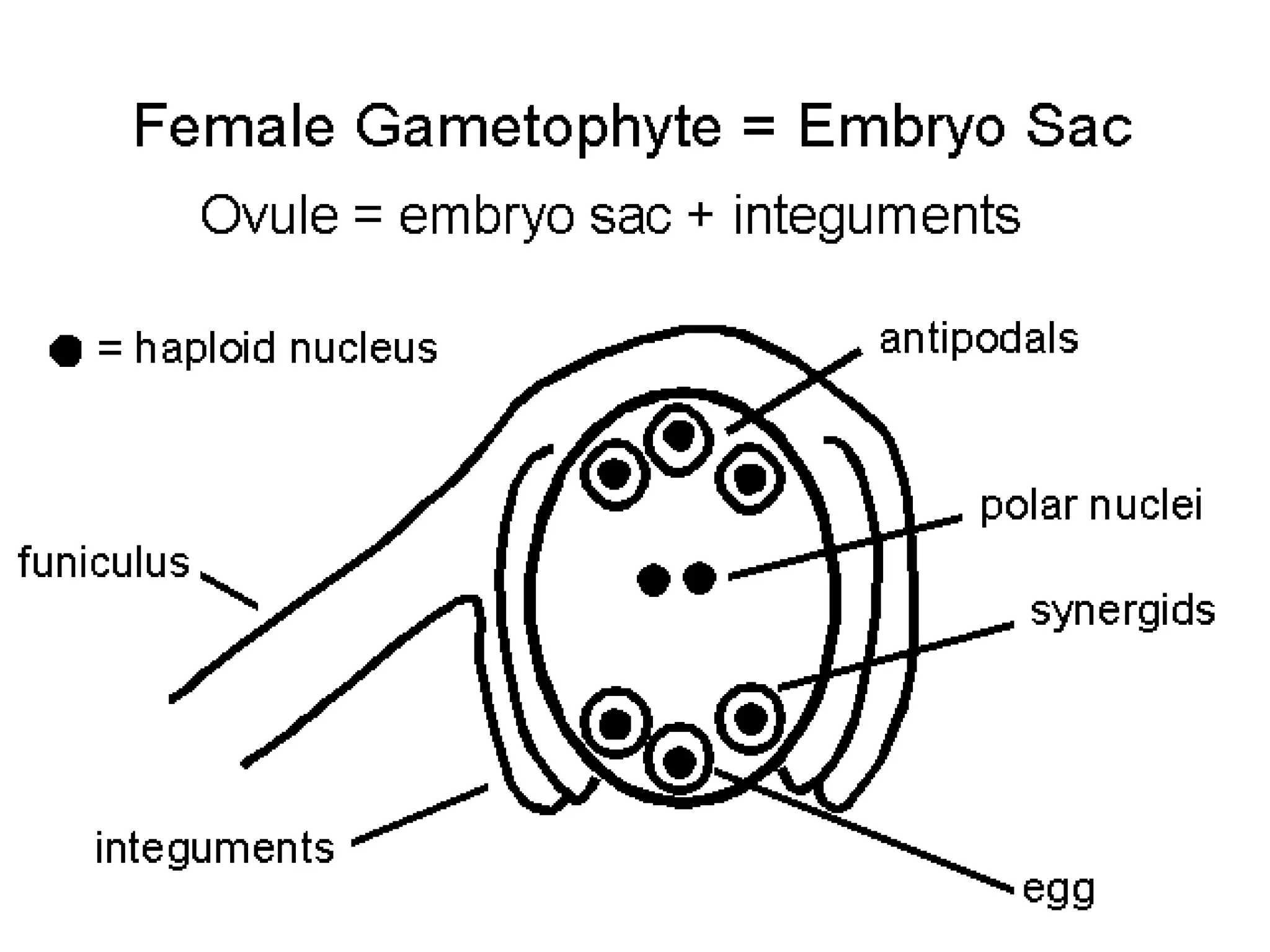
The document details the structure and development of ovules within the gynoecium, outlining key components such as pistils, stigma, style, ovary, integuments, and the nucellus. It explains the processes of megasporogenesis and the formation of female gametophytes, highlighting the roles of various structures like micropyle, obturator, and raphe in reproduction. Additional information includes variations in integument structures and the evolution of megasporangium characteristics across different plant families.