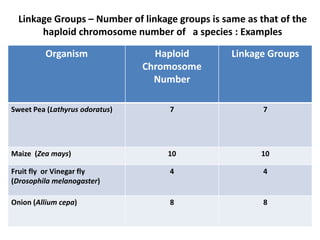
The document discusses the concepts of linkage and crossing-over in genetics, particularly through the example of sweet pea and Drosophila melanogaster experiments. It explains how the experiments of Bateson, Saunders, and Punnett led to the development of the coupling and repulsion theory, which was later refined by Morgan's theory of linkage, highlighting the importance of gene proximity on inheritance patterns. Additionally, it outlines the differences between complete and incomplete linkage, providing examples of how they affect phenotypic ratios in offspring.



![• British geneticists, Bateson,
Saunders and Punnett (1906)
had carried out hybridization
experiments in two varieties of
Sweet pea (Lathyrus odoratus).
• In this experiment, they had
focused the inheritance of two
pairs of contrasting characters
such as flower colour (purple
vs. red) and shape of pollen
grains (long vs. round), [a
dihybrid cross]. One Sweet
pea plant was with purple
flowers and long pollen grains
and the other with red flowers
and round pollen grains.
Sweet pea
(Lathyrus odoratus)
Edith Rebecca Saunders, William Bateson &
Reginald Crundall Punnett
Creative Commons Attribution 4.0 International ,https://www.researchgate.net , © 2014 Cornell University,
https://pollen.tstebler.ch
Pollen grains of
Sweet pea](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-3-320.jpg)
![• Let ‘P’ stands for the allele for purple flower colour and ‘L’ , for the
allele for long pollen grains. So, the genotype (genetic constitution)
of the pure breeding homozygous Sweet pea parent plant with
purple flowers and long pollen grains can be represented as PPLL .
• From previous hybridization experiments in Sweet pea, it had been
found that, purple flower colour is dominant over red and long
pollen grain is dominant over round. Hence the genotype of the
other Sweet pea plant with red flowers and round pollen grains
(both recessive traits) can be represented as ppll .
• All the F1 (First filial generation) plants had purple flowers and long
pollen grains (hybrids, Genotype in heterozygous condition, PpLl) as
expected.
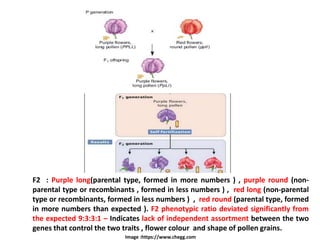
• On further crossing (selfing, F1 x F1 – PpLl x PpLl ), the F1 plants
produced the F2 generation (Second filial generation).
[Alleles : Two or more alternative forms of the same gene having different
phenotypic effects]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-4-320.jpg)




![The double heterozygote , PpLl in
Coupling Phase
(PPLL x ppll → PpLl – Coupling double heterozygote)
Here, the dominant alleles, P and L are on one chromosome and their recessive
alleles p and l are on the other chromosome of a homologous pair – i.e., in
Coupling phase.
[Homologous pair : The maternal and paternal chromosomes]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-10-320.jpg)











![Example : Complete Linkage in male Drosophila melanogaster
• Morgan and Lynch (1911) had found that in Drosophila, the
genes for body colour (gray vs. black) and wing size (long vs.
vestigial ) were linked to each other.
• It is also to be noted that the allele for gray body colour is
dominant over black and the allele for long wings is dominant
over vestigial wings.
Gray body, long wings Black body , vestigial wings
https://sciencemusicvideos.com
[Long wings : Normal functional wings
Vestigial wings : Shriveled or crumpled, non-functional wings ]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-23-320.jpg)


![• In test cross, non-parentals or recombinants (flies with gray
body , vestigial wings and black body , long wings) were not
produced because of complete linkage between the genes
for gray body colour and long wings and between black body
colour and vestigial wings in the F1 double heterozygote
male fly so that they remained together during inheritance
(GL/gl). Hence, there is lack of independent assortment
and crossing-over.
[Test cross to detect linkage: Test cross (Hybrid x homozygous recessive
parent) can be used to detect linkage. A deviation from the typical test
cross ratio (where both parental phenotypes and recombinant phenotypes
are formed in equal proportions) is an indication of linkage].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-26-320.jpg)
![Complete Linkage in male Drosophila melanogaster
In this figure, the chromosomes are drawn as rod shaped .The results of the test cross between F1 male Drosophila
fly ( GgLl – can be represented as GL/gl in coupling or cis linkage phase, Gray long) with double recessive female
parent fly (ggll, Black vestigial) deviated from the expected dihybrid test cross ratio, 1:1:1:1(1 gray long [parentals] :
1 gray vestigial [recombinants] : 1 black long [recombinants] : 1 black vestigial [parentals] ). Instead , the progeny
include only parentals, Gray long and Black vestigial , produced in equal proportions. That is, 50 % Gray long : 50%
Black vestigial i.e. in a ratio, 1:1 and no recombinants are produced . This is due to complete linkage and hence
absence of crossing-over in F1 male Drosophila fly.](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-27-320.jpg)

![Example: Incomplete Linkage in female Drosophila melanogaster
• Calvin Bridges crossed a wild type female Drosophila fly with
gray body and long wings (Genotype GGLL) with a male
having black body and vestigial wings (Genotype ggll).
• The F1 flies were wild type with gray body and long wings
(Genotype GgLl, can be represented as GL/gl in coupling or
cis linkage phase).
• A test cross between F1 female fly and the double recessive
male parent fly produced progeny with more parental types
(gray long and black vestigial, 83 %) and a few non-parental
types or recombinants (gray vestigial and black long, 17 %)
and thus deviated from the typical dihybrid test cross
ratio,1: 1: 1: 1, where both parental types and recombinants
are produced in equal proportions(i.e., 1 gray long : 1 gray
vestigial : 1 black long : 1 black vestigial).
[ Wild type : The most common phenotype in a natural population].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-29-320.jpg)




![• Linkage analysis is an effective method to locate a gene of
interest in a chromosome . Linkage analysis has helped to
find out the location of several genes on human
chromosomes and to study the genetic basis of many
human diseases.
• Linkage restrict genetic variability in a population.
[Genetic variability is essential for hereditary improvement of traits].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-34-320.jpg)

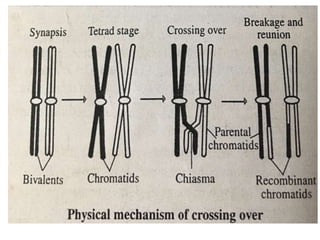
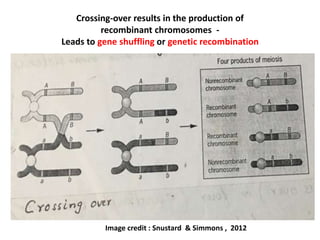
![• Crossing-over involves the reciprocal
exchange of equivalent portions
between non-sister chromatids of a pair
of homologous chromosomes. Morgan
and Cattell (1912) had called this process
of interchanging, "crossing-over".
• The crossing-over break the linkage
between genes that leads to shuffling
and genetic recombination. Therefore,
crossing-over results in new
combination of alleles in the gametes.
• That means, crossing-over alters the
pattern of genes in the chromosomes
and thus creates genetic variability
among population.
[Population : A group of organisms of one
species that interbreed and live in the same
place at the same time].
Image :https://www.toppr.com](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-36-320.jpg)

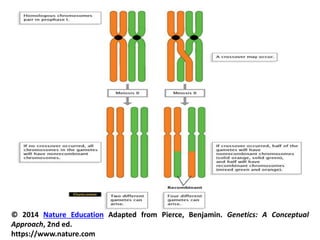
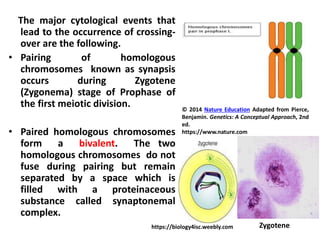
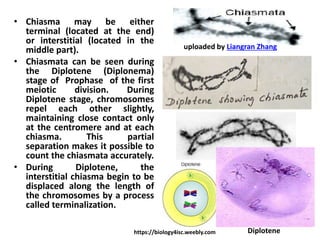
![• During the next stage Pachytene (Pachynema), the
chromosomes become shorter and thicker. Each
chromosome split length wise (divide) to form two
chromatids (sister chromatids), held together by the
undivided centromere. Thus , the bivalent consists
of four chromatids (four strand stage or tetrad).
Crossing-over occurs at this stage.
• Crossing-over takes place by symmetrical breakage,
reciprocal exchange and reattachment of the pieces
between any two non-sister chromatids of the
tetrad.
[Double-strand breaks: Programmed double-strand
breaks by an endonuclease, Spo11(DNA
topoisomerase VI subunit A) at specific chromosomal
positions into segments – Formation of double-
strand breaks does not occur uniformly across the
genome. There are certain regions which are
favorable for double-strand break formation known
as double-strand break hotspots such as intergenic
and/or promoter regions where nucleosomes are
more widely dispersed . The union of segments of
chromosomes is catalysed by an enzyme , DNA
ligase.
Hotspots : Location of some intense activity]
© 2014 Nature Education Adapted
from Pierce, Benjamin. Genetics: A
Conceptual Approach, 2nd ed.
https://www.nature.com
https://biology4isc.weebly.com
Pachytene](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-40-320.jpg)

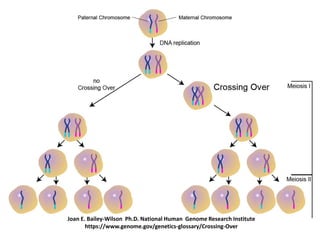


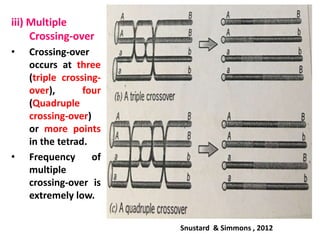

![• The number of crossing-over is strictly regulated to form an optimum
number of physical links (i.e., chiasmata) between homologous
chromosomes to ensure their accurate segregation during anaphase
of meiosis I.
• Even if there is fluctuation in DNA double-strand break formation, the
frequency of crossing-over is maintained more or less constant
(Crossover homeostasis).
• The formation of crossing-over is regulated in such a way that their
number is kept at a very low level. This may help to maintain
the DNA sequences in cells with very little change from generation to
generation. In most of the species, the average number of crossing-
over per chromosome rarely exceeds three per bivalent.
• However, several genetic, epigenetic (= relating to or arising from
non-genetic influences on gene expression e.g. DNA methylation) and
environmental factors can influence the frequency of crossing-over.
[DNA methylation : Addition of methyl (CH3) group from S-adenyl methionine to
the fifth carbon (C5 position) of a cytosine to form 5-methylcytosine in DNA.
Methylation can change the activity of a segment of DNA without changing the
sequence. DNA methylation play a role in epigenetic gene regulation.]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-52-320.jpg)


![• The percentage of crossing-over (recombination) or Cross over value can be
calculated as
Where, total number of progeny = Total No. of parental types + Total No.
of recombinants.
(As already stated, when there is random assortment (i.e. independent assortment)
of genes, the parental types and the recombinants will be formed in equal
proportions. That means in a 1:1 ratio).
On substitution, we get
1/(1+ 1) x 100 = 50%.
[i.e. 1(Total No. of Recombinants) / 1(Total No. of parental types) + 1 (Total
No. of recombinants ) x 100].
• Thus, the recombination frequency between two independently assorting
genes cannot exceed 50%. In other words, the maximum recombination
frequency between two independently assorting genes is 50%.](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-55-320.jpg)
![• Sturtevant(1914), had suggested that frequency of recombination could be
used to estimate the relative distance between two genes. 1% Recombination
frequency = 1 map unit = 1 cM. In other words, 1 cM indicates a 1%
recombination potential. That means, in 100 meioses, there will be one
recombination event .
• Thus, two genes with a low recombination frequency are likely to be closer
together while those with a high recombination frequency are likely to be
farther apart on a chromosome.
• Based on this, Sturtevant created the first genetic map ( Linkage map) of
genes on the X chromosome of Drosophila melanogaster.
[ cM – centiMorgan,(in honor of Thomas Hunt Morgan), the unit of map
distance.
Map distance : According to Sturtevant(1914), percentage of crossing-over is
used as an index of the distance between any two pairs of genes. That is, the
unit of distance is taken as a portion of the chromosome of such length that, on
an average, crossing-over occurs in it in 1% of the germ cells(reproductive cells).
i.e., one map unit is equal to 1% recombinant phenotypes. In other words, a
spacing of 1 cM indicates a 1 % chance that two genes will be separated by
crossing-over ].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-56-320.jpg)
![ Crossover Assurance, Interference and Coincidence
Crossover Assurance
• Irrespective of size, at least one crossing-over (obligate crossing-over)
occurs per bivalent, generating the ‘obligate chiasma’ that ensures
proper disjunction of homologous chromosomes during anaphase I
of Meiosis I. This phenomenon is called ‘Crossover Assurance’ .
Failure to maintain at least one crossing-over per homologous pair of
chromosomes increases the probability of non-disjunction which in
turn may result in the formation of aneuploid gametes.
[Disjunction : Normal separation of chromosomes toward opposite poles
during anaphase.
Non-disjunction : The failure of one or more pairs of homologous
chromosomes or sister chromatids to separate normally during nuclear
division, usually resulting in an abnormal distribution of chromosomes in the
daughter nuclei.
Aneuploidy : The loss or gain of one or a few chromosomes].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-57-320.jpg)


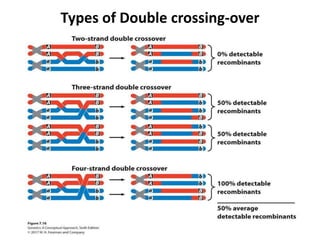
![Coincidence
• The simultaneous occurrence of two single crossing-over events in
the same pair of homologous chromosomes to form a double
crossing-over is called coincidence .
• Coefficient of coincidence (CoC ) = Observed frequency of double
crossovers / Expected frequency of double crossovers in target
intervals .
[ In other words, Number of double recombinants in the progeny /
Number of double recombinants expected in the progeny ;
Expected double crossovers can be calculated as
Expected double crossovers = Recombination frequency in region 1
X Recombination frequency in region 2. i.e., the probability of a
double crossover is the product of their separate probabilities].
• Coefficient of Coincidence (CoC) + Interference (I) = 1
Therefore, interference , I can be calculated as
I = 1 – CoC .](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-60-320.jpg)


![ Position of gene
• The correlation between distance between genes and the probability of
crossing-over may not be applicable to all genes in a chromosome. This is
because, formation of chiasma does not occur at random throughout the
length of a chromosome.
• Crossing-over occur in preferential regions of the genome. That means,
certain regions of some chromosomes have significantly higher rates of
crossing-over or recombination called crossing-over hotspots or
recombination hotspots, which serve as localized stimulators of general
recombination and other regions which have unusually low recombination
called coldspots, which depress the level of recombination. Recombination
hotspots are interspersed with recombination coldspots.
[Crossing-over hotspots: DNA fragments of a few kilobases in length with a higher rate of
recombination than the surrounding DNA - Hotspots occur in very small regions, approximately
25% of the genome, where 80% of the crossing-over occur. For instance, in many primates, mice
etc., 80 % of recombination occur in 10-20 % of the genome. In humans, recombination hotspots
are approximately 10% of the genome where 40-60 % recombination occur. In many plant
species also, recombination events are not evenly distributed along the length of the
chromosomes. For instance, in wheat (Triticum aestivum L.) chromosome 3B, it has been
estimated that 90% of the crossing-over occur in distal sub-telomeric regions. In Arabidopsis,
crossing-over hotspots are closely associated with DNA double-strand break hotspots (Choi et. al.,
2018) ].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-63-320.jpg)


![ Chromatin Structure
• Frequency of crossing-over can be influenced by chromatin
structure. In Maize, frequency of crossing-over is more in the distal
gene-rich euchromatic regions.
• Crossing-over can be suppressed by methylation of DNA and
histones ; the amount and distribution of highly repetitive
heterochromatin etc. The occurrence of crossing-over is rare or
absent in heterochromatic regions. Genomic regions with
repetitive sequences have low frequency of crossing-over.
• Recombination frequency is positively correlated with G-C content
in certain organisms like Drosophila melanogaster, Saccharomyces
cerevisiae ( brewer's yeast or baker's yeast), honey bee etc. There
are also reports that regions with low G-C content have high
recombination (Lynn et.al., 2004). But, in Arabidopsis thaliana,
there is no correlation between G-C content and the frequency of
crossing-over.
[ G-for Guanine and C-for Cytosine, the nitrogen bases in DNA]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-66-320.jpg)
![• Recombination frequency is also influenced by transposable
elements. Generally, crossing-over is suppressed in highly
repetitive genomic regions that are made up of transposable
elements (Underwood and Choi, 2019).
• Crossing-over hotspots identified from Maize have low DNA
methylation and transposons. High transposable element
densities have been reported in low recombination regions of
Drosophila species. On the other hand, Theobroma cacao and rice
populations show largely divergent hotspot locations influenced
by retrotransposon abundance and genetic divergence (Marand
et. al., 2019).
[ Transposable elements (Transposons or Jumping genes) : They can move
from one location on the genome to another - first discovered by Barbara
Mc Clintock in Maize .
Retrotransposon : They transpose through RNA intermediates. First, the
transposable DNA is copied into RNA and then into DNA by reverse
transcription by an enzyme reverse transcriptase and inserted into the
target site]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-67-320.jpg)
![• Generally, there is a positive correlation between gene density and
frequency of crossing-over.
• Formation of crossing-over is affected by sequence homology. For
instance, in the case of inversion heterozygotes and translocation
heterozygotes, where one chromosome has the normal sequence and
the other chromosome with altered sequence, the pairing of
homologous chromosomes will be disrupted in the regions of
chromosomes where such changes have occurred , which will in turn
reduce the probability of crossing-over. That is, reduction in sequence
homology between chromosomes will lessen the probability of crossing-
over and thereby recombination.
[Gene density : The number of genes per million base pairs, called a
megabase, Mb ; For instance, the gene density of the human genome is
roughly 12–15 genes/Mb.]
Inversion : A type of structural change in a chromosome in which two
breaks occur and the broken segment is reinserted after rotating 180° so
that the gene order is reversed.
Translocation: A type of chromosomal aberration which involves the
transfer of a segment of a chromosome to a different part of the same
chromosome or to a different chromosome, which change the
arrangement of the genes ].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-68-320.jpg)
![ Autopolyploidy
• Autopolyploids generally exhibit reduced crossing-over rates
when compared with their diploids (Shaver, 1962; Watanabe,
1983; Gillies et al., 1987; Yant et al., 2013).
Sex of the individual
• In most of the organisms, crossing-over occur in both males and
females. But, there are exceptions. Crossing-over is absent in male
Drosophila melanogaster (Morgan, 1914 ; but Drosophila
ananassae males undergo a few crossing-over )and female silk
moth Bombyx mori (Tanaka, 1914) ( Achiasmy – Complete
suppression of recombination in one sex).
• Sex-specific differences in the frequency of crossing-over
(Heterochiasmy) and its distribution along chromosomes have
been reported in many species.
[Autopolyploidy : The phenomenon where all the genomes in a polyploid
organism are identical
Polyploid : An organism with more than two sets of chromosomes].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-69-320.jpg)
![• Frequency of crossing-over may be
a) Higher in female meiosis – E.g. Eutherian mammals - For
instance, crossing-over frequency in oocytes of human females is
generally higher than in spermatocytes of human males,
which correlates with differences in synaptonemal complex length -
The synaptonemal complex is considerably longer in oocytes in
comparison to spermatocytes. (Tease and Hultén, 2004 ; Petkov
et.al., 2007).
b) Higher in male meiosis - In some metatherian mammals, Sheep,
Arabidopsis thaliana etc.
In the plant Arabidopsis thaliana, crossing-over rates in distal
regions of chromosome 4 are very high in male meiosis but very
low in female meiosis ( Drouaud et.al., 2007).
c) Almost same - There is no significant sex-specific differences in
the rate of crossing-over. e.g. Tomato, Barley, Rape seed or Canola
etc.
[Eutherian mammals : Placental mammals that give birth to well-developed
young ones.
Metatherian mammals : They give birth to partially developed young ones e.g.
Marsupiales].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-70-320.jpg)



![• In Arabidopsis thaliana, both low (8°C) and high (28°C)
temperatures (Cold and heat stresses), increase the frequency
of crossing-over. An increase in temperature within the
fertility-tolerable range (28°C) promotes crossing-over. For
instance, frequency of crossing-over increased when
Arabidopsis plant grown at a temperature of 20°C was
shifted to 28°C. The frequency of crossing-over was
approximately 10% higher at the extreme of the
temperature range, 8–28 °C.
• An increase in chromosome axis length may account for the
rise in frequency of crossing-over at low temperature in
Arabidopsis thaliana (Lloyd et. al., 2018).
• A higher temperature (32°C - 38°C) disrupts central element
of synaptonemal complex and causes asynapsis and thus
affects bivalent and chiasma formation which in turn result
in the suppression of crossing-over (De Storme and Geelen,
2020).
[Asynapsis : Failure of pairing of homologous chromosomes]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-74-320.jpg)




![ Plasma genes (Cytoplasmic Genes)
• In some species, plasma genes may cause reduction in
crossing-over. For example, Tifton male sterile cytoplasm
(Tift 23 A1 cytoplasm) in Pearl millet (Bajra - Pennisetum
glaucum).
[Tift 23 A : Cytoplasmic-genic male sterile (CMS) line of Pearl millet with
short stature, profuse tillering, uniform flowering and good combining
ability, evolved at Tifton, Georgia. In Pearl millet , the first reported CMS
system, A1 was based on the Tift 23A1 cytoplasm (Burton, 1965, Burton
and Athwal 1967) ; used in commercial hybrid seed production in Pearl
millet.]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-79-320.jpg)
![ Radiations
• Investigations on effect of radiation on frequency of crossing-over gave
conflicting results. The different responses of organisms to radiation may be
due to various factors such as biological ( i.e., species-specific differences
and variation in the developmental stage during which irradiation was done)
and physical ( i.e., nature of the radiation employed and the temperature
when irradiation was done).
• Generally, there was a decrease in the frequency of crossing-over after
irradiation.
• Plants irradiated just before the start of meiosis had much lower chiasma
frequency .
• Generally, crossing-over is absent in male Drosophila flies. But, extremely
low frequency of crossing-over had been reported in male Drosophila flies
(Muller, 1916; Bridges and Morgan, 1919 ; Sturtevant, 1929 ; Patterson and
Suche, 1933). It has been found that crossing-over can be induced in male
Drosophila flies by X-ray irradiation of immature germ cells. Rifenburgh
(1935) had reported that irradiation of young larvae by ultra-violet radiation
induced crossing-over between the black and vestigial loci in Drosophila
male fly.
[Irradiation : Exposure of a biological material to any one of the radiations ]](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-80-320.jpg)

![Significance of Crossing-over
1. Creation of Genetic variability
• Crossing-over and genetic recombination in the first meiotic
division during gametogenesis is an essential feature of sexual
reproduction that increase genetic variability among the
progeny, which is essential for effective selection (both
natural and artificial). Genetic variability is a prerequisite for
the evolutionary process.
• However, the low number of crossing-overs often limits the
genetic variation that can be utilized in breeding
programs(Plant Breeding and Animal husbandry).
[Plant Breeding : Applied branch of Botany that deals with the genetic
improvement of crops for the service of man.
Animal husbandry : The science of breeding and caring of domesticated animals].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-82-320.jpg)




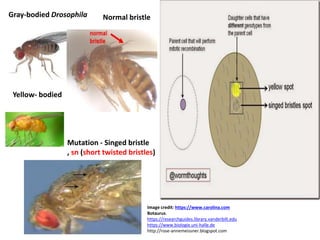
![Mitotic Crossing-over
• Crossing-over that occurs in somatic cells (i.e., body cells)
during mitosis (i.e., somatic crossing-over). It is a rare event
that occurs with a frequency of 10-4 to 10-5 per cell division
(Gunther, 1984).
• Mitotic crossing-over was first reported by Curt Stern (1936) on
X chromosome of a female Drosophila melanogaster fly with
the heterozygous genotype +sn/y+ (Repulsion or Trans linkage
phase) for body colour (wild type, gray ‘+’ vs. yellow, ‘y’) and
bristles ( normal ‘+’ vs. singed ‘sn’ – gnarled or short twisted
bristles). That is, one X chromosome carry the recessive allele
‘y’ for yellow body and the other X chromosome carry the
recessive allele ‘sn’ for singed bristles.
[Female Drosophila fly has two X chromosomes. i.e., XX].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-87-320.jpg)
![• Stern had observed that most of the female Drosophila flies were
gray bodied and with normal bristles, as gray body colour is
dominant over yellow and normal bristles is dominant over
singed. However, Stern had also observed that some female flies
had single yellow spots, singed spots and twin yellow-singed
spots(i.e.,twin sectors, yellow adjacent to singed) on gray body.
This is because of mitotic crossing-over and segregation that
occured in a cell with heterozygous genotype, +sn/y+, which
results in cells homozygous for y and for sn. Thus mitotic
recombination resulted in the expression of recessive genes in
small areas in wild type gray bodied female fly.
[Twin spots : Paired alterations visible in adjacent areas – mosaic patches or
spots.
Organisms that are composed of cells of more than one genotype are referred
to as genetic mosaics].](https://image.slidesharecdn.com/linkageandcrossingover-220524140908-9381e277/85/Linkage-and-Crossing-over-pptx-88-320.jpg)